![]()
![]()
![]()
LES PECHERIES PELAGIQUES
Les espèces de thon ayant une importance commerciale sont en Mauritanie au nombre de trois:
l'albacore (Thunnus albacares),
le listao (Katsuwonus pelamis),
le patudo (Thunnus obesus).
Ces espèces sont exploitées par une flottille de canneurs basée à Dakar, et pêchant sous licence, dans les eaux mauritaniennes. La saison de pêche débute selon les années en mai-juin, et se poursuit jusqu'en novembre-décembre. L'abondance des thons dans la ZEE mauritanienne est très liée à la présence des eaux chaudes au large de la Mauritanie. Les captures de thons en Mauritanie varient selon les années entre 5 et 10000 tonnes pour un potentiel estimé à 10–14000 tonnes (Marcille, 1986).
L'activité des flottes de pêche industrielle est dirigée sur un nombre limité d'espèces cibles appartenant aux groupes suivants:
les carangidés avec les chinchards noirs (Trachurus trachurus et T. trecae) en proportion variable suivant les années ou les secteurs, et le chinchard jaune (Decapterus rhonchus);
les clupéidés avec la sardine (Sardina pilchardus), les sardinelles plate (Sardinella maderensis) et ronde (S. aurita) avec généralement une prédominance de cette dernière;
les scombridés représentés surtout par le maquereau espagnol (Scomber japonicus) qui ne correspond à une espèce cible qu'en des périodes limitées.
Les prises accessories réalisées lors des pêches dirigées sont composées soit d'espèces à comportement pélagique dominant comme l'anchois (Engraulis encrasicolus) ou surtout le poisson sabre (Trichiurus lepturus), ou le calmar, soit d'espèces davantage inféodées au fond. Parmi ces dernières on trouve principalement le pelon (Brachydeuterus auritus), les merlus, et divers sparidés.
Les espèces cibles exploitées par les flottilles de pêche industrielle sont caractérisées par leur comportement migratoire qui les amène à entreprendre d'importants déplacements (Garcia, 1982) en fonction des variations hydroclimatiques caractérisées par les mouvements saisonniers du front thermique (cf. Chapitre I et annexe C).
Les ressources pélagiques de la ZEE mauritanienne sont exploitées par des flottilles de senneurs et de chalutiers dont la situation (effectif et effort) depuis 1979 a été revue.
La flottille internationale “Interpêche” a cessé toute activité en 1981 (Josse, 1987). Depuis mai 1988, six senneurs portugais sont autorisés, dans le cadre des accords CEE, à travailler dans la ZEEM. Leurs caractéristiques sont les suivantes:
| nb | LT(m) | TJB | équipage |
|---|---|---|---|
| 2 | 14 à 17 | 20 à 31 | 9 |
| 4 | 29 à 35 | 140 à 200 | 18 à 24 |
Ils constituent une flotte industrielle à long rayon d'action travaillant dans le cadre de sociétés mixtes ou sous licences. Depuis 1984 (date des données les plus récentes analysées par le premier Groupe de travail) les principaux changements concernent l'arrêt de l'activité des chalutiers fournisseurs de la RDA (juin 83), celui de la flotte bulgare (août 83), cubaine (jullet 85), irakienne (septembre 86), allemande de l'est (juin 87). Par ailleurs, les chalutiers du Nigéria n'ont fréquenté la ZEE mauritanienne qu'entre septembre 1985 et octobre 1986 et les soviétiques de la classe des STM ont été présents d'octobre 1984 à juin 1987. Les effectifs correspondants, avec les caractéristiques de jauge et de puissance de chaque classe de navire sont donnés, par nationalité dans le tableau IV.1.
En raison de la pratique des remplacements entre chalutiers qui permet à des navires différents de se succéder sur zone, le nombre de chalutiers effectivement présents dans la ZEE ne correspond pas au nombre d'autorisations délivrées; ce dernier est donné, à titre de comparaison dans le tableau IV.2 pour les saisons de pêche 1986/1987, 1987/1988 et 1988/1989.
Les eaux situées en deçà des 3 milles de la ligne de base cap Blanc - cap Timiris et en deçà des 3 milles de la côte au sud de Timiris étaient interdites aux senneurs de la flottille “Interpêche”. La nouvelle réglementation appliquée aux senneurs nationaux en opération depuis la mi-88 interdit l'accès des eaux côtières situées à moins de 6 milles de la ligne de base cap Blanc - cap Timiris ou de la côte au sud de ce dernier.
La réglementation a été modifiée à partir du ler juillet 1987. Avant cette date, les chalutiers des flottilles de pêche industrielle étaient maintenus au delà des 12 milles de la ligne de base cap Blanc - cap Timiris ou de la côte au sud de ce cap (Figure IV.1a). Depuis lors, la zone d'interdiction a été repoussée, au niveau de la région cap Blanc - cap Timiris, au delà d'une ligne suivant approximativement la sonde des 30 mètres (Figure IV.1b).
Aucune donnée nouvelle n'est disponible. L'analyse de l'effort déployé par la flottille de senneurs jusqu'en 1981 est disponible dans le Bulletin du CNROP n° 15.1 (Josse, 1987). Les seules données détaillées concement la flottille “Interpêche” pour les années 1980 et 1981. Pour le reste les données ne couvrent que quelques mois et/ou certaine(s) flottille(s).
L'effort de pêche global développé par les flottilles des différentes nationalités a été réévalué pour la période 1979–1987 (Tableau IV.3). Ces efforts sont exprimés en jour de pêche et par classe de navire. Bien que l'effort de pêche des flottes soviétiques soit quantitativement le plus important il ne peut servir de base, dans l'état actuel des possibilités de traitement et d'analyse, au calcul des prises par unité d'effort (P.U.E.) en raison de l'irrégularité de leur présence (sauf pour les trois dernières années) et de la forte saisonnalité des pêcheries. Par contre, malgré la disparition d'une composante de la flotte roumaine (fin 83) l'effort développé par cette dernière parait suffisamment régulier et fiable pour être utilisé dans le calcul d'une P.U.E.
| 1979 | 1980 | 1981 | 1982 | 1983 | 1984 | 1985 | 1986 | 1987 | |
|---|---|---|---|---|---|---|---|---|---|
| Allemagne de l'Est | |||||||||
| 995 TJB - 1750 cv | 0 | 13 | 13 | 13 | 12 | 0 | 0 | 0 | 0 |
| 3165 TJB - 3880 cv | 5 | 5 | 5 | 5 | 5 | 2 | 3 | 2 | 3 |
| Bulgarie | |||||||||
| 2500 TJB - 3600 cv | 0 | 0 | 0 | 4 | 5 | 0 | 0 | 0 | 0 |
| Cuba | |||||||||
| 2635 TJB - 4000 cv | 0 | 0 | 0 | 0 | 0 | 7 | 4 | 0 | 0 |
| Irak | |||||||||
| 2326 TJB - 2000 cv | 0 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 0 |
| 2406 TJB - 3600 cv | 0 | 0 | 3 | 4 | 4 | 5 | 4 | 3 | 0 |
| Nigéria | |||||||||
| 535 TJB - 2000 cv | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 5 | 0 |
| 1000 TJB - 3600 cv | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Roumanie | |||||||||
| 2156 TJB - 2320 cv | 5 | 5 | 3 | 4 | 3 | 0 | 0 | 0 | 0 |
| 3163 TJB - 3880 cv | 2 | 10 | 13 | 11 | 9 | 10 | 11 | 12 | 17 |
| URSS | |||||||||
| 1900 TJB - 2400 cv | 0 | 0 | 0 | 0 | 0 | 2 | 10 | 4 | 2 |
| 2400 TJB - 2000 cv | 0 | 7 | 0 | 5 | 8 | 17 | 18 | 13 | 12 |
| 2177 TJB - 2320 cv | 0 | 2 | 0 | 3 | 20 | 17 | 24 | 15 | 14 |
| 3019 TJB - 3880 cv | 0 | 10 | 0 | 16 | 38 | 23 | 20 | 19 | 11 |
| 2900 - 3400 TJB - 5600 cv | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 |
| 1986/1987 | 1987/1988 | 1988/1989 | |
|---|---|---|---|
| Allemagne de l'Est | |||
| 3165 TJB – 3880 cv | 2 | 2 | 0 |
| Irak | |||
| 2326 TJB – 2000 cv | 4 | 0 | 0 |
| 2406 TJB – 3600 cv | 3 | 0 | 0 |
| Roumanie | |||
| 3163 TJB – 3880 cv | 8 | 13 | 12 |
| URSS | |||
| 1900 TJB – 2400 cv | 3 | 0 | 0 |
| 2400 TJB – 2000 cv | 9 | 3 | 4 |
| 2177 TJB – 2320 cv | 11 | 14 | 13 |
| 3019 TJB – 3880 cv | 13 | 10 | 10 |
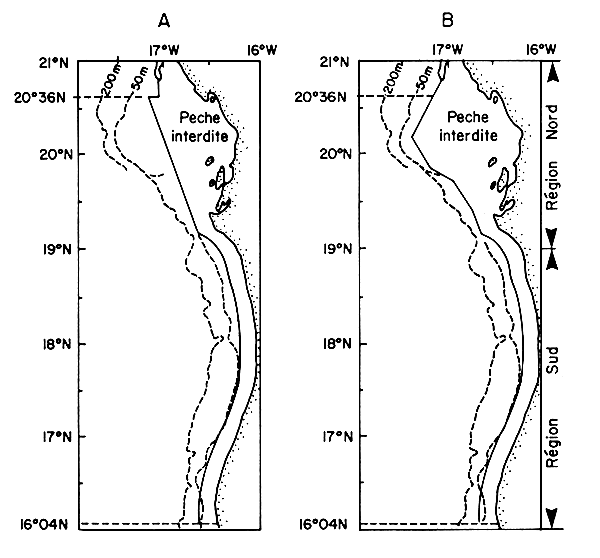
Figure IV.1. - Réglementation des zones de pêches concernant les chalutiers pélagiques industriels (d'après Chavance, 1988).
| 1979 | 1980 | 1981 | 1982 | 1983 | 1984 | 1985 | 1986 | 1987 | |
|---|---|---|---|---|---|---|---|---|---|
| Allemagne de l'Est | |||||||||
| 995 TJB - 1750 cv | 0 | 1599 | 2133 | 1973 | 964 | 0 | 0 | 0 | 0 |
| 3165 TJB - 3880 cv | 553 | 795 | 1224 | 1296 | 996 | 631 | 432 | 341 | 472 |
| Bulgarie | |||||||||
| 2500 TJB - 3600 cv | 0 | 0 | 0 | 314 | 392 | 0 | 0 | 0 | 0 |
| Cuba | |||||||||
| 2635 TJB - 4000 cv | 0 | 0 | 0 | 0 | 0 | 563 | 318 | 0 | 0 |
| Irak | |||||||||
| 2326 TJB - 2000 cv | 0 | 80 | 960 | 720 | 937 | 894 | 1079 | 655 | 0 |
| 2406 TJB - 3600 cv | 0 | 0 | 660 | 660 | 660 | 628 | 682 | 282 | 0 |
| Nigéria | |||||||||
| 535 TJB - 2000 cv | 0 | 0 | 0 | 0 | 0 | 0 | 149 | 1022 | 0 |
| 1000 TJB - 3600 cv | 0 | 0 | 51 | 0 | 0 | 0 | 0 | 0 | 0 |
| Roumanie | |||||||||
| 2156 TJB - 2320 cv | 519 | 410 | 468 | 518 | 0 | 0 | 0 | 0 | |
| 3163 TJB - 3880 cv | 974 | 1323 | 1965 | 2300 | 1469 | 1316 | 2002 | 1855 | 2772 |
| URSS | |||||||||
| 1900 TJB - 2400 cv | 0 | 0 | 0 | 0 | 0 | 115 | 651 | 372 | 164 |
| 2400 TJB - 2000 cv | 0 | 547 | 0 | 237 | 1271 | 1222 | 2064 | 2145 | 1782 |
| 2177 TJB - 2320 cv | 0 | 162 | 0 | 100 | 2530 | 1215 | 2418 | 2142 | 2815 |
| 3019 TJB - 3880 | 0 | 798 | 0 | 957 | 4976 | 2513 | 2434 | 2601 | 2170 |
| 2900 à 3400 TJB - 5600 cv | 0 | 0 | 0 | 49 | 216 | 0 | 0 | 0 | 0 |
| TOTAL | 1527 | 5823 | 7403 | 9074 | 14929 | 9097 | 12229 | 11415 | 10175 |
L'examen des variations de l'effort de pêche roumain depuis 1972 (Figure IV.2) doit tenir compte du fait que jusqu'en 1978 (inclus) la fréquentation de la zone 34.1.3 intéressait essentiellement les eaux situées au large du Sahara occidental. Après 1979 qui constituait une année intermédiaire, l'effort attribué à la zone 34.1.3 s'est reporté en totalité vers la ZEE mauritanienne. Le niveau de fréquentation est alors sensiblement identique dans les deux zones sauf en 1987 où il s'est porté vers le nord (34.1.3) où il a atteint 71%. Ce comportement inhabituel s'oppose à celui de la flotte soviétique qui de son côté et contrairement aux années précédentes, a reporté une fraction importante de son effort vers la zone méridionale (34.3.1).
Les captures des chalutiers et des senneurs par espèce et groupe d'espèces ont été également révisées pour la période 1979–1987 (Tableau IV.4).
Leur capture a culminé en 1980 avec 281 000 tonnes dont 45% était composé de sardinelles.
La capture globale des espèces pélagiques s'est accrue régulièrement au cours de la période récente pour atteindre 445 000 t environ en 1987. Elle est largement dominée par les chinchards qui représentent environ 51% de cette capture. Les productions par espèce ou groupe d'espèces pour les chalutiers seuls ou pour les deux métiers combinés (senne et chalut) sont données dans les figures IV.3 et IV.4 respectivement.
La distribution géographique des captures (Chavance, 1988) permet d'identifier deux secteurs principaux, l'un en région nord où 31 % des pêches est effectué dans seulement deux carrés statistiques de 20' de côté. Par contre, dans la région méridionale, les pêches sont plus dispersées; moins de 24 % des prises y est effectué dans quatre carrés statistiques (Figure IV.5). En période chaude (juillet-septembre), les flottilles fréquentent surtout le secteur nord; elles commencent leur descente très progressivement vers le secteur sud en octobre. Cette dernière est achevée au mois de mai. En l'espace d'un mois (juin), les flottilles exécutent une remontée massive vers le secteur nord (Figure IV.6).
Des séries de P.U.E. ont été établies à partir des données de captures et d'effort de la flotte roumaine (Tableau IV.5). La P.U.E. pour les chinchards montre une brusque et importante augmentation vers la fin des années 70 pour décroître tout aussi rapidement au cours des années 80 (Figure IV.7). Cette fluctuation parait être en opposition de phase avec celle de la P.U.E. calculée pour les clupéidés totaux même si les valeurs absolues et l'importance des variations de cette dernière sont moindres. Pour les trois dernières années, les mêmes indices concernant la flotte soviétique sont également indiqués.
En détaillant la fluctuation des P.U.E. des clupéidés pour la sardine d'une part et les sardinelles d'autre part (Figure IV.8), la première espèce est bien responsable de l'oscillation observée au cours des années 70 et les sardinelles ont engendré une seconde oscillation au cours des années 80. De plus, d'après les données roumaines une augmentation sensible de la P.U.E. de sardine semble se dessiner depuis 1985 quoique cette tendance s'inverse brutalement entre 1986 et 1987 dans la série d'indices soviétiques. On se souviendra cependant que la flotte soviétique a porté en 1987 une part importante de son effort dans la zone méridionale.
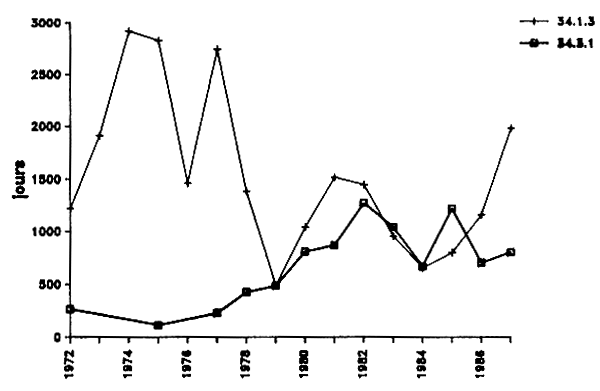
Figure IV.2. - Evolution de l'effort de pêche roumain (exprimé en nombre de jours de pêche) pour les zones COPACE 34.1.3 et 34.3.1 de 1972 à 1987 (Origine des données : Maxim et Staicu, 1987 ; Staicu et Maxim, 1984, 1987 ; Staicu et al, 1988).
| Type d'exploitation | 1979 | 1980 | 1981 | 1982 Chalut | 1983 Chalut | 1984 Chalut | 1985 Chalut | 1996 Chalut | 1987 Chalut | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Senne* | Chalut | Senne* | Chalut | Senne* | Chalut | |||||||
| Sardine | 25438 | 3387 | 56037 | 14961 | 7933 | 0 | 0 | 72939 | 36099 | 51490 | 81925 | 34792 |
| Sardinelles | 63936 | 2358 | 125340 | 14443 | 91806 | 24082 | 42136 | 64509 | 66331 | 94047 | 61457 | 65309 |
| Chinchards | 24730 | 71353 | 93131 | 190132 | 10340 | 176079 | 151961 | 196943 | 176929 | 199617 | 205902 | 228093 |
| Maquereau | 0 | 9387 | 0 | 10895 | 597 | 12108 | 10108 | 46400 | 13459 | 17482 | 28800 | 24982 |
| Autres | 3546 | 915 | 20 | 14050 | 0 | 19604 | 70503 | 89940 | 68020 | 91994 | 78939 | 91919 |
| TT pélagique | 117650 | 87400 | 274528 | 244481 | 11676 | 213873 | 274782 | 470731 | 337947 | 454630 | 457023 | 445095 |
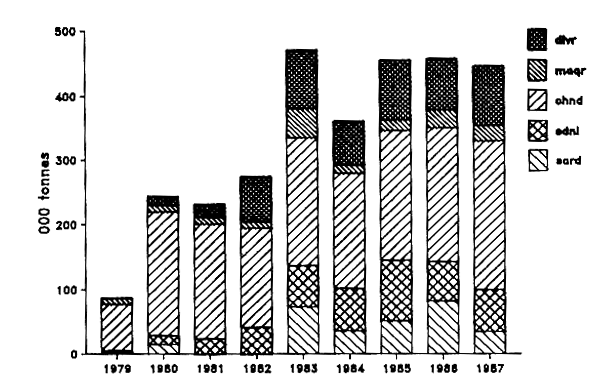
Figure IV.3. - Captures par espèces, ou groupes d'espèces, des chalutiers pélagiques industriels dans la ZEE mauritanienne de 1979 à 1987 (Origine des données : tableau IV.4).
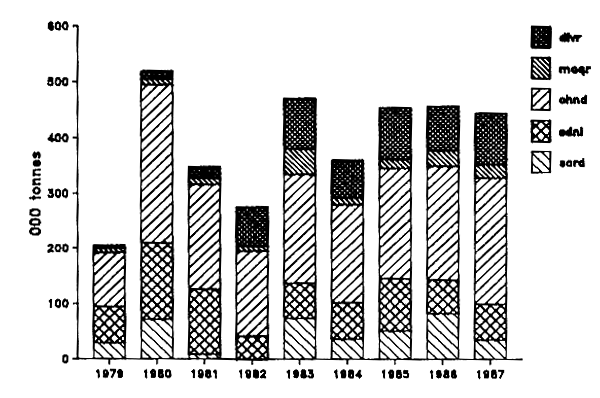
Figure IV.4. - CAptures par espèces de la pêche pélagique industrielle dans la ZEE mauritanienne de 1979 à 1987 (Origine des données : tableau IV.4).
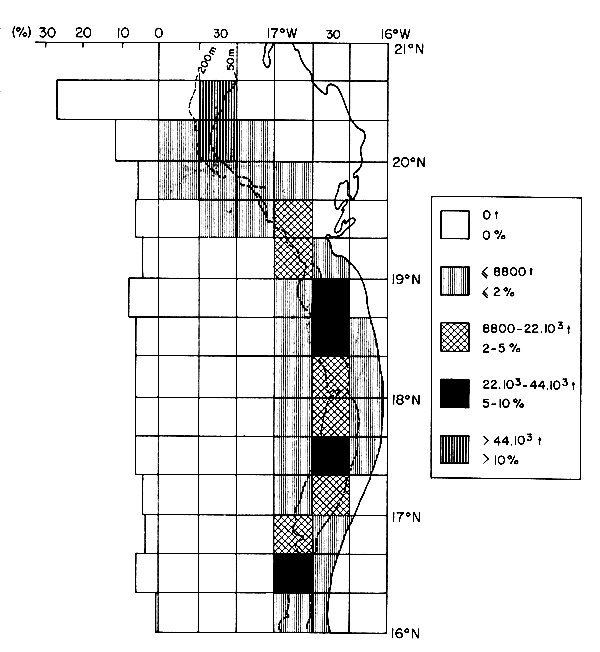
Figure IV.5. - Répartition spatiale, par “carrés” statiques de 20 minutes de côté des captures de cha;itoers pélagiques industriels dans la ZEE mauritanienne en 1987 (d'après Chavance, 1988).
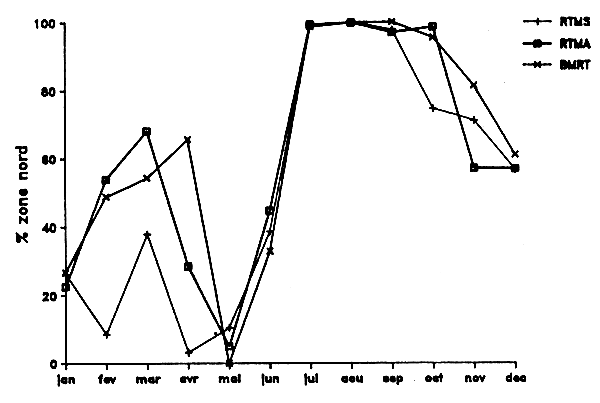
Figure IV.6. - Répartition nord-sud de l'effort des flottilles soviétiques de chalutiers pélagiques dans la ZEE mauritanienne en 1987 (Origine des données : statistiques officielles de l'AtlanNIRO).
| Année | 1972 | 1973 | 1974 | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 | 1981 | 1982 | 1983 | 1984 | 1985 | 1986 | 1987 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Effort de pêche | ||||||||||||||||
| 34.1.3. (nord) | 1219 | 1909 | 2914 | 2818 | 1457 | 2733 | 1379 | 486 | 1036 | 1509 | 1439 | 948 | 649 | 792 | 1154 | 1971 |
| 34.3.1. (sud) | 268 | 114 | 230 | 428 | 488 | 806 | 866 | 1269 | 1039 | 667 | 1215 | 701 | 801 | |||
| Total en jours | 1487 | 1909 | 2914 | 2932 | 1457 | 2963 | 1807 | 974 | 1842 | 2375 | 2708 | 1987 | 1316 | 2007 | 1855 | 2772 |
| Captures ( t ) | ||||||||||||||||
| chinchards | 19576 | 30477 | 39267 | 29491 | 14144 | 39190 | 33832 | 41488 | 64214 | 64099 | 44537 | 20235 | 17739 | 45409 | 45775 | 71232 |
| sardine | 0 | 122 | 21275 | 34939 | 17641 | 26968 | 10566 | 678 | 4541 | 0 | 0 | 2510 | 0 | 2716 | 8425 | 16808 |
| sardinelles | 3303 | 4110 | 1188 | 1821 | 611 | 1523 | 960 | 272 | 3137 | 8872 | 16039 | 18495 | 13781 | 22321 | 15593 | 16593 |
| clupéidés | 3303 | 4232 | 22463 | 36760 | 18252 | 28491 | 11526 | 950 | 7678 | 8872 | 16039 | 21005 | 13781 | 25037 | 24018 | 33401 |
| P.u.e. (t/j) | ||||||||||||||||
| chinchards | 13.165 | 15.965 | 13.475 | 10.058 | 9.7076 | 13.226 | 18.723 | 42.595 | 34.861 | 26.989 | 16.446 | 10.184 | 13.479 | 22.625 | 24.677 | 25.697 |
| sardine | 0 | .06391 | 7.3010 | 11.916 | 12.108 | 9.1016 | 5.8473 | .69610 | 2.4653 | 0 | 0 | 1.2632 | 0 | 1.3533 | 4.5418 | 6.0635 |
| sardinelles | 2.2213 | 2.1530 | .40769 | .62108 | .41935 | .51401 | .53127 | .27926 | 1.7030 | 3.7356 | 5.9228 | 9.3080 | 10.472 | 11.122 | 8.4059 | 5.9859 |
| clupéidés | 2.2213 | 2.2169 | 7.7086 | 12.538 | 12.527 | 9.6156 | 6.3785 | .97536 | 4.1683 | 3.7356 | 5.9228 | 10.571 | 10.472 | 12.475 | 12.948 | 12.049 |
Origine des données: Staicu et Maxim 1984 Maxim et Staicu 1987 Staicu et Maxim 1987 Staicu et al in Anonyme 1988
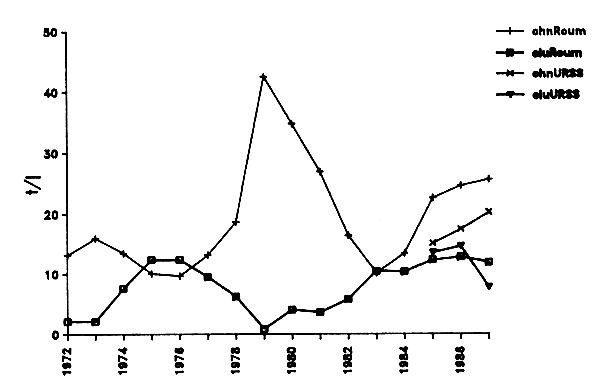
Figure IV.7. - Evolution des P.U.E. (en tonnes/jour de pêche) “Chinchards” et “Clupéidés” pour les flottilles roumaines (1972–1987) et soviétiques (1985–1987). Origine des données : tableau IV. 5 et statistiques officielles de l'ATlanNIRO).
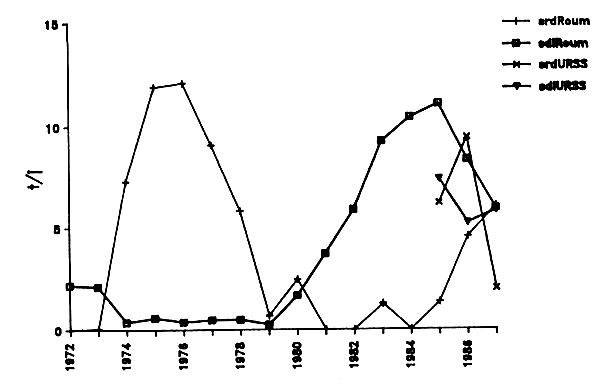
Figure IV.8. - Evolution des P.U.E. (en tonnes/jour de pêche) “Chinchards” et “Clupéidés” pour les flottilles roumaines (1972–1987) et soviétiques (1985–1987). Origine des données : tableau IV. 5 et statistiques officielles de l'ATlanNIRO).
Ce résumé reprend l'inventaire effectué lors du dernier Groupe de travail CNROP-CRODT/ISRA sur les Ressources Pélagiques (Anonyme, 1988). Il est complété par:
les résultats récemment acquis sur l'anchois(Annexe I),
les données portant sur les coefficients de mortalité disponibles dans la littérature,
les coefficients de mortalité naturelle déduits pour les principales espèces pélagiques de l'équation de Pauly (1980).
Ba I., 1988 - Biologie et dynamique des populations d'anchois (Engraulis encrasicolus) des côtes mauritaniennes. Thèse de 3ème cycle. Université de Bretagne Occidentale, Brest : 138 p.
Barkova N.A. et Domanevsky L.N., 1986 - Etat des stocks de la sardine (Sardina pilchardus Walb.) dans la division statistique 34.1.3 COPACE/PACE Séries 86/39: 80–96.
Belvèze H., 1984 - La pêcherie de maquereaux (Scomber japonicus) de l'Atlantique marocain entre 29°N et 33°N. COPACE/PACE Séries 83/27: 104–127.
Boély T. 1979 - Biologie des deux espèces de sardinelles (Sardinella aurita Valenciennes 1847 et Sardinella maderensis Lowe 1841) des côtes sénégalaises. Thèse de doctorat d'état. Université Pierre et Marie Curie, Paris VI. 219 p.
Boély T., Fréon P. et Stéquert B., 1982 - La croissance de Sardinella aurita (val. 1847) au Sénégal. Océanogr. Trop., 17(2): 103–119.
Boély T., Wysokinsky A. et Elwertowski J., 1973 - les chinchards des côtes sénégalaises. Doc. Sci.provis.Centr.Rech. Océanogr.Dakar-Thiaroye, ORSTOM 46:47 p.
Camarena T.L., 1986 - Les principales espèces de poissons pélagiques côtiers au Sénégal: Biologie et évaluations des Ressources. Thèse de 3ème Cycle. Université de Bretagne Occidentale, Brest: 187 p.
Conand F. et Franqueville C., 1973 - Identification et distribution saisonnière de larves de carangidés au large du Sénégal et de la Gambie. Bull. I.F.A.N.XXXV, Sér.A, n°4: 953–978.
El Ouairi M., 1984 - Quelques données sur le stock du chinchard de l'Atlantique marocain. COPACE/PACE Séries 83/27: 129–134.
FAO, Comité des Pêches pour l'Atlantique Centre-est3, 1979(a) - Rapport du Groupe de travail ad hoc sur les poissions pélagiques côtiers ouest-africains de la Mauritanie au Libéria (26°N à 5°N). COPACE/PACE Séries 78/10 : 165 p.
FAO, Comité des Pêches pour l'Atlantique Centre-est, 1979 (b)- Rapport du séminaire pour l'unification des techniques de lecture d'âge et d'interprétation de Sardina pilchardus. COPACE/TECH 78/18: 23 p.
3 Voir également Fréon et al, Krivospitchenko S.G., Overko S.M. et Sedletskaia V.A., 1979
| ANNEE | PARAMETRE | PARAMETRE | UNITE | UNITE | GAMME DE TAILLE | ZONES | SOURCES | NATIONALITE |
|---|---|---|---|---|---|---|---|---|
| a | b | P | L | |||||
| Sardina pilchardus | ||||||||
| 1973 | 0.055 | 2.536 | GR | LFCM | 18–23 | 34.1.3 | FAO,1986(p105) | ROUMANIE |
| 1974 | 0.018 | 2.824 | GR | LFCM | 11–22 | 34.1.3 | FAO,1986(p105) | ROUMANIE |
| 1975 | 0.009 | 3.098 | GR | LFCM | 12–22 | 34.1.3 | FAO,1986(p105) | ROUMANIE |
| 1976 | 0.021 | 2.827 | GR | LFCM | 15–23 | 34.1.3 | FAO,1986(p105) | ROUMANIE |
| 1977 | 0.009 | 3.106 | GR | LFCM | 13–22 | 34.1.3 | FAO,1986(p105) | ROUMANIE |
| 1978 | 0.009 | 3.114 | GR | LFCM | 16–23 | 34.1.3 | FAO,1986(p105) | ROUMANIE |
| 1983 | 0.016 | 2.91 | GR | LFCM | 18–23 | MAURITANIE | FAO,1986(p105) | ROUMANIE |
| 1976–77 | 0.0075 | 3.24 | GR | LFCM | 7–19 | SENEGAL | FREON ET STEQUERT,1979 | SENEGAL |
| 1978–88 | 0.01881 | 2.8837 | GR | LFCM | 13–22 | MAURITANIE | LAWAL ET MYLNIKOV,1988 | MAURITANIE |
| Sardinella aurita | ||||||||
| 1968–69 males | 0.0000018 | 3.388 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| femelles | 0.0000019 | 3.3768 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| indet | 0.0000032 | 3.2817 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| tot | 0.0000052 | 3.1943 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| 1968(1°trim) | 0.0000034 | 3.2816 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| 1968(2°trim) | 0.0000023 | 3.3363 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| 1968(3°trim) | 0.0000052 | 3.1953 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| 1968(4°trim) | 0.0000000 | 3.0342 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| 1969(1°trim) | 0.0000022 | 3.3506 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| 1969(2°trim) | 0.0000038 | 3.2583 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| 1969(3°trim) | 0.0000003 | 2.813 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| 1969(4°trim) | 0.0000074 | 3.1123 | GR | LFMM | SENEGAL | BOELY,1979 | SENEGAL | |
| 1976–77 | 0.006392 | 3.274 | GR | LFCM | 4–32 | SENEGAL | FAO,1979(p122) | SENEGAL |
| ? | 0.01648 | 2.8386 | GR | LTCM | AFRIQUE DE L'OUEST | FAO,1979(p7) | ||
| 1981 | 0.0024471 | 3.3751 | GR | LTCM | 22–37 | MAURITANIE | HOLZLOHNER ET KLOXIN,1982 | RDA |
| 1982(1°trim) | 0.004 | 3.8854 | GR | LTCM | 31–40 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(2°trim) | 0.0038 | 3.2607 | GR | LTCM | 27–38 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(3°trim) | 0.01 | 2.9725 | GR | LTCM | 26–40 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(4°trim) | 0.003 | 3.3398 | GR | LTCM | 16–36 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1984–85 | 0.0061104 | 3.2901 | GR | LFCM | 5–32 | SENEGAL | CAMARENA,1986 | SENEGAL |
| 1987–88 | 0.00794 | 3.2278 | GR | LFCM | 15–32 | MAURITANIE | LAWAL ET MYLNIKOV, 1988 | MAURITANIE |
| Sardinella maderensis | ||||||||
| 1975–77? | 0.01034 | 3.142 | GR | LFCM | 4–29 | SENEGAL | FAO,1979(p123) | SENEGAL |
| 1982(1°trim) | 0.018 | 2.8328 | GR | LTCM | 32–38 | MAURITANIE | HOLZLOHNER ET AL, 1983 | RDA |
| 1982(2°trim) | 0.0264 | 2.7202 | GR | LTCM | 28–38 | MAURITANIE | HOLZLOHNER ET AL, 1983 | RDA |
| 1982(3°trim) | 0.0053 | 3.163 | GR | LTCM | 25–37 | MAURITANIE | HOLZLOHNER ET AL, 1983 | RDA |
| 1984–85 | 0.0098535 | 3.1673 | GR | LFCM | 5–29 | SENEGAL | CAMARENA,1986 | SENEGAL |
| 1985 | 0.0229297 | 2.778069 | GR | LFCM | 7–29 | CAP VERT | FAO, 1987(p150) | CAP VERT |
| 1987–88 | 0.0098 | 3.183 | GR | LFCM | 8–30 | MAURITANIE | LAWAL ET MYLNIKOV, 1988 | MAURITANIE |
| Engraulis encrasicolus | ||||||||
| 1986–1987 | 0.00147 | 3,75 | GR | LFCM | 6,5–13,0 | MAURITANIE | BA I..1988 | MAURITANIE |
| ANNEE | PARAMETRE | PARAMETRE | UNITE | UNITE | GAMME DE TAILLE | ZONES | SOURCES | NATIONALITE |
|---|---|---|---|---|---|---|---|---|
| a | b | P | L | |||||
| Trachurus trachurus | ||||||||
| 1978–79 | 0.019 | 2.876 | GR | ?CM | >15 CM | MAROC | FAO,1984(p130) | MAROC |
| ? | 0.023 | 2.505 | GR | ?CM | <10 CM | MAROC | FAO,1984(p130) | MAROC |
| ? | 0.005 | 3.12 | GR | ?CM | >10 CM | MAROC | FAO,1984(p130) | MAROC |
| 1973 | 0.0197 | 2.85 | GR | LFCM | 18–34 | 20–26°N | FAO,1984(p53) | ROUMANIE |
| 1974 | 0.0106 | 3.05 | GR | LFCM | 6–34 | 20–26°N | FAO,1984(p53) | ROUMANIE |
| 1975 | 0.0207 | 2.85 | GR | LFCM | 9–32 | 20–26°N | FAO,1984(p53) | ROUMANIE |
| 1976 | 0.0089 | 3.09 | GR | LFCM | 9–35 | 20–25°N | FAO,1984(p53) | ROUMANIE |
| 1977 | 0.0292 | 2.73 | GR | LFCM | 10–32 | 17–29°N | FAO,1984(p53) | ROUMANIE |
| 1978 | 0.0208 | 2.83 | GR | LFCM | 17–35 | 16–24°N | FAO,1984(p53) | ROUMANIE |
| 1979 | 0.0446 | 2.6 | GR | LFCM | 14–36 | 16–23°N | FAO,1984(p53) | ROUMANIE |
| 1980 | 0.0125 | 2.98 | GR | LFCM | 12–34 | MAURITANIE | FAO,1984(p53) | ROUMANIE |
| 1981 | 0.0198 | 2.85 | GR | LFCM | 17–33 | MAURITANIE | FAO,1984(p53) | ROUMANIE |
| 1982 | 0.0128 | 2.99 | GR | LFCM | 10–37 | MAURITANIE | FAO,1984(p53) | ROUMANIE |
| 1981 | 0.0265 | 2.685 | GR | LTCM | 25–43 | MAURITANIE | FAO,1984(p53) | RDA |
| 1982(1°TRIM) | 0.0087 | 2.2995 | GR | LTCM | 27–40 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(2°TRIM) | 0.0388 | 2.5943 | GR | LTCM | 31–40 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(3°TRIM) | 0.0023 | 3.3803 | GR | LTCM | 21–37 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(4°TRIM) | 0.0096 | 3.1484 | GR | LTCM | 25–41 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| ? | 0.01 | 3.02 | GR | LFCM | 14–36 | 14–26°N | FAO,1984(p3) | URSS |
| ? | 0.0049 | 3.14 | GR | LTCM | ? | ? | FAO,1979(p9) | POLOGNE |
| 1981 | 0.027 | 2.6936 | GR | LTCM | 20–45 | MAURITANIE | HOFFMANN,1987 | RDA |
| 1982 | 0.0545 | 2.4785 | GR | LTCM | 7–40 | MAURITANIE | HOFFMANN,1987 | RDA |
| 1983 | 0.0128 | 2.9986 | GR | LFCM | 17–35 | MAURITANIE | HOFFMANN,1987 | RDA |
| 1984 | 0.015 | 2.9364 | GR | LFCM | 14–38 | MAURITANIE | HOFFMANN,1987 | RDA |
| 1987–88 | 0.008 | 3.147 | GR | LFCM | 8–35 | MAURITANIE | LAWAL ET MYLNIKOV,1988 | MAURITANIE |
| Trachurus trecae | ||||||||
| 1976? | 0.1489 | 2.954 | GR | LFCM | 7–40 | SENEGAL | FAO,1979(p126) | SENEGAL |
| 1977? | 0.01 | 2.98 | GR | LFCM | 8–43 | FAO,1984(p7) | URSS | |
| 1973 | 0.0247 | 2.8 | GR | LFCM | 16–32 | 20–26°N | FAO,1984(p53) | ROUMANIE |
| 1974 | 0.0314 | 2.72 | GR | LFCM | 12–33 | 20–26°N | FAO,1984(p53) | ROUMANIE |
| 1975 | 0.0145 | 2.96 | GR | LFCM | 18–31 | 20–26°N | FAO,1984(p53) | ROUMANIE |
| 1976 | 0.0101 | 3.07 | GR | LFCM | 15–30 | 20–25°N | FAO,1984(p53) | ROUMANIE |
| 1981 | 0.0083 | 3.11 | GR | LFCM | 9–30 | MAURITANIE | FAO,1984(p53) | ROUMANIE |
| 1982 | 0.0248 | 2.8 | GR | LFCM | 14–35 | MAURITANIE | FAO,1984(p53) | ROUMANIE |
| 1982 | 0.0248344 | 2.809017 | GR | LFCM | 14–36 | MAURITANIE | FAO,1984(p7) | ROUMANIE |
| 1981 | 0.0106 | 2.9366 | GR | LTCM | 14–44 | MAURITANIE | FAO,1984(p7) | RDA |
| 1982(1°trim) | 0.0062 | 3.0612 | GR | LTCM | 19–37 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(2°trim) | 0.025 | 2.7171 | GR | LTCM | 24–44 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(3°trim) | 0.0057 | 3.1232 | GR | LTCM | 21–45 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(4°trim) | 0.0138 | 2.8722 | GR | LTCM | 19–49 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1984–85 | 0.01569 | 3.0422 | GR | LFCM | 9–34 | SENEGAL | CAMARENA,1986 | SENEGAL |
| 1987–88 | 0.011 | 3.041 | GR | LFCM | 4–41 | MAURITANIE | LAWAL ET MYLNIKOV,1988 | MAURITANIE |
| ANNEE | PARAMETRE | PARAMETRE | UNITE | UNITE | GAMME DE TAILLE | ZONES | SOURCES | NATIONALITE |
|---|---|---|---|---|---|---|---|---|
| a | b | P | L | |||||
| Decapterus rhonchus | ||||||||
| 1972–1974 | 0.0799232 | 2.50429 | GR | LFCM | 22–40 | 17–26°N | MAXIM ET STAICU,1976 | ROUMANIE |
| 1975–1976 | 0.01243 | 3.055 | GR | LFCM | 4–41 | SENEGAL | FAO,1984(p11) | SENEGAL |
| ? | 0.006521 | 3.097 | GR | LTCM | ? | ? | FAO,1979(p12) | POLOGNE |
| 1981 | 0.0125 | 2.9092 | GR | LTCM | 26–45 | MAURITANIE | FAO,1984(p10) | RDA |
| ? | 0.02 | 2.88 | GR | LFCM | 15–40 | SAHARA-MAURIT. | FAO,1984(p11) | URSS |
| 1982(1*trim) | 0.0204 | 2.7991 | GR | LTCM | 21–30 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(2*trim) | 0.0083 | 3.0286 | GR | LTCM | 13–33 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(3*trim) | 0.0145 | 2.8545 | GR | LTCM | 17–33 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(4*trim) | 0.0237 | 2.7351 | GR | LTCM | 15–33 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1980 | 0.0149 | 2.8838 | GR | LTCM | 26–44 | MAURITANIE | HOFFMANN,1987 | RDA |
| 1981 | 0.0129 | 2.9145 | GR | LTCM | 26–49 | MAURITANIE | HOFFMANN,1987 | RDA |
| 1982 | 0.0209 | 2.7769 | GR | LTCM | 22–45 | MAURITANIE | HOFFMANN,1987 | RDA |
| 1983 | 0.008 | 3.1656 | GR | LFCM | 17–39 | MAURITANIE | HOFFMANN,1987 | RDA |
| 1984 | 0.0336 | 2.7467 | GR | LFCM | 23–37 | MAURITANIE | HOFFMANN,1987 | RDA |
| 1985 | 0.0142 | 2.9647 | GR | LFCM | 7–38 | MAURITANIE | *MIMEO,1987 | RDA |
| 1986 | 0.01233 | 3.0608 | GR | LFCM | 7–38 | SENEGAL | CAMARENA,1986 | SENEGAL |
| 1987–88 | 0.021 | 2.91 | GR | LFCM | 19–38 | MAURITANIE | LAWAL ET MYLNIKOV,1988 | MAURITANIE |
| *présenté au groupe de travail mais non publié | ||||||||
| Scomber japonicus | ||||||||
| 1970–73 | 0.0019355 | 3.567567 | GR | LFCM | 14–43 | 12–29°N | FAO,1984(p14) | ROUMANIE |
| 1975–82 | 0.00407 | 3.3 | GR | LF?CM | 9–39 | MAROC | FOA,1984(p110) | MAROC |
| 1978? | 0.004871 | 3.278 | GR | LFCM | 8–41 | SENEGAL | FAO,1979(p124) | SENEGAL |
| 1981 | 0.0018 | 3.4744 | GR | LTCM | 26–48 | MAURITANIE | HOLZLOHNER ET KLOXIN,1982 | RDA |
| 1982(2°trim) | 0.0022 | 3.437 | GR | LTCM | 32–42 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(3°trim) | 0.0006 | 3.8071 | GR | LTCM | 26–36 | MAURITANIE | HOLZLOHNER ET AL,1983 | RDA |
| 1982(4°trim) | 0.0014 | 3.5573 | GR | LTCM | 25–48 | MAURTIANIE | HOLZLOHNER ET AL,1983 | RDA |
| ? | 0.003916 | 3.352 | GR | LFCM | ?-53 | SAH-MAURIT-SEN | FAO,1984(p14) | URSS |
| 1984–85 | 0.003892 | 3.3619 | GR | LFCM | 6–41 | SENEGAL | CAMARENA,1986 | SENEGAL |
| 1987–88 | 0.000189 | 3.577 | GR | LFCM | 11–44 | MAURITANIE | LAWAL ET MYLNIKOV,1988 | MAURITANIE |
| ANNEE | Linfini | K | t0 | GAMME DE TAILLE (cm) | ZONES | METHODES | SOURCES | NATIONALITE |
|---|---|---|---|---|---|---|---|---|
| Sardina pilchardus | ||||||||
| ? | 21.63 | 0.49 | -1.07 | ? | MAROC | ECAILLES | FAO,1979(b,p18) | MAROC |
| ? | 22.96 | 0.517 | -0.84 | ? | SAHARA | OTOLITHES | FAO,1979(b,p18) | POLOGNE |
| ?males | 22.96 | 0.522 | -0.68 | ? | SAHARA | OTOLITHES | FAO,1979(b,p18) | POLOGNE |
| ?femelles | 23.52 | 0.482 | -0.95 | ? | SAHARA | OTOLITHES | FAO,1979(b,p18) | POLOGNE |
| ? | 24.62 | 0.371 | -1.32 | ? | SAHARA | OTOLITHES | FAO,1979(b,p18) | POLOGNE |
| ?femelles | 24.53 | 0.381 | -1.35 | ? | SAHARA | OTOLITHES | FAO,1979(b,p18) | POLOGNE |
| ?males | 23.16 | 0.437 | -1.12 | ? | SAHARA | OTOLITHES | FAO,1979(b,p18) | POLOGNE |
| ? | 21.23 | 0.68 | -1.33 | ? | MAROC | ECAILLES | FAO,1979(b,p18) | MAROC |
| ? | 20.27 | 0.981 | 0.02 | ? | MAROC | ECAILLES | FAO,1979(b,p18) | MAROC |
| ? | 20.25 | 0.926 | 0.06 | ? | MAROC | ECAILLES | FAO,1979(b,p18) | MAROC |
| ?femelles | 20.46 | 0.943 | 0.08 | ? | MAROC | ECAILLES | FAO,1979(b,p18) | MAROC |
| ?males | 20.06 | 0.906 | 0.03 | ? | MAROC | ECAILLES | FAO,1979(b,p18) | ? |
| ? | 22.81 | 0.946 | 0.22 | ? | CAP BLANC | ECAILLES | FAO,1979(b,p18) | ? |
| ? | 21.58 | 0.82 | -1.29 | ? | MAROC | OTOLITHES | FAO,1979(b,p18) | ? |
| ? | 21.93 | 0.8 | -0.38 | ? | MAROC | ECAILLES | FAO,1979(b,p18) | ? |
| ?males | 21.67 | 0.8 | -0.27 | ? | MAROC | ECAILLES | FAO,1979(b,p18) | ? |
| ?femelles | 22.02 | 0.82 | -0.34 | ? | MAROC | ECAILLES | FAO,1979(b,p18) | ? |
| 1973 | 22.8 | 0.51 | -0.205 | 18-LF-23 | 34.1.3 | ECAILLES | FAO,1986(p105) | ROUMANIE |
| 1974 | 23.08 | 0.42 | -0.95 | 11-LF-22 | 34.1.3 | ECAILLES | FAO,1986(p105) | ROUMANIE |
| 1975 | 22.17 | 0.49 | -0.826 | 12-LF-22 | 34.1.3 | ECAILLES | FAO,1986(p105) | ROUMANIE |
| 1976 | 22.49 | 0.43 | -0.954 | 15-LF-22 | 34.1.3 | ECAILLES | FAO,1986(p105) | ROUMANIE |
| 1977 | 22.56 | 0.39 | -1.48 | 13-LF-22 | 34.1.3 | ECAILLES | FAO,1986(p105) | ROUMANIE |
| 1978 | 22.16 | 0.42 | -1.402 | 16-LF-23 | 34.1.3 | ECAILLES | FAO,1986(p105) | ROUMANIE |
| 1983 | 23.98 | 0.33 | -1.523 | 18-LF-23 | 34.1.3 | ECAILLES | FAO,1986(p105) | ROUMANIE |
| 1981–83 | 25.6 | 048 | -0.39 | 9-LF-24 | 34.1.3 | ? | FAO,1986(p94) | URSS |
| Sardinella aurita | ||||||||
| 1966–77 | 30.63 | 1.206 | -0.062 | 6-LF-32 | SENEGAL | FREQ.DE TAILLE | BOELY ET AL,1982 | SENEGAL |
| ET ECAILLES | ||||||||
| 1984–85 | 31.93 | 0.613 | -0.745 | 17-LF-32 | SENEGAL | ECAILLES | CAMARENA,1986 | SENEGAL |
| ? | 40.7 | 0.326 | -0.6283 | LT | MAURITANIE | ? | PHAM THUOC ET SZYPULA,1973 | ? |
| Sardinella maderensis | ||||||||
| 1980–82 | 39.5 | 0.45 | 9-LF-30 | SENEGAL | FREQ.DE TAILLE | SAMB,1986 | SENEGAL | |
| 1980–82 | 37.5 | 0.3 | 9-LF-30 | SENEGAL | FREQ.DE TAILLE | SAMB,1986 | SENEGAL | |
| (wp=0,1 | c=0,6)* | * equation de croissance saisonnière | ||||||
| 1984–85 | 30.34 | 0.49 | -0.589 | 8-LF-30 | SENEGAL | FREQ.DE TAILLE | CAMARENA,1986 | SENEGAL |
| ET ECAILLES | ||||||||
| Engraulis encrasicolus | ||||||||
| 1986–1987 | 21.8 | 1.056 | 0.0675 | 4-LF-13,5 | MAURITANIE | OTOLITHES | BA I.,1988 | MAURITANIE |
| MARQUES JOURNALIERES | ||||||||
| ANNEE | Linfini | K | t0 | GAMME DE TAILLE (cm) | ZONES | METHODES | SOURCES | NATIONALITE |
|---|---|---|---|---|---|---|---|---|
| Trachurus trachurus | ||||||||
| ? | 37.98 | 0.25 | -0.98 | LF | 21–26°N | ECAILLES | FAO,1979(p9) | URSS |
| ? | 50 | 0.13 | -2.32 | LT | ? | OTOLITHES | FAO,1979(p9) | POLOGNE |
| ? | 43.55 | 0.19 | -1.33 | LF | 34.1.1 | ? | FAO,1984(p5) | URSS |
| ? | 40.88 | 0.22 | -1.31 | LF | 34.1.3 | ? | FAO,1984(p5) | URSS |
| 1972–82 | 38.98 | 0.2775724 | -1.1613564 | LF | SEN-MAURITANIE | ? | FAO,1984(p5) | ROUMANIE |
| ? | 61.36 | 0.1011 | -0.85 | ? | MAROC | OTOLITHES | FAO,1984(p129) | MAROC |
| ? | 53.3 | 0.12 | -0.3 | ? | MAROC | OTOLITHES | FAO,1984(p130) | MAROC |
| 1978–79 | 43.39 | 0.19 | -1.31 | ? | MAROC | ? | FAO,1984(p130) | MAROC |
| 1981 | 60.03 | 0.0854 | -3.734 | 25-LT-42 | MAURITANIE | OTOLITHES | HOFFMANN,1987 | RDA |
| 1982 | 46.34 | 0.1877 | -1.463 | 20-LT-41 | MAURITANIE | OTOLITHES | HOFFMANN,1987 | RDA |
| 1983 | 36.06 | 0.357 | -0.475 | 17-LF-39 | MAURITANIE | OTOLITHES | HOFFMANN,1987 | RDA |
| 1984 | 45.82 | 0.2006 | -1.02 | 14-LF-39 | MAURITANIE | OTOLITHES | HOFFMANN,1987 | RDA |
| 1982(1°trim) | 51.863 | 0.1574 | -1.61 | 27-LT-40 | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL,1983 | RDA |
| 1982(3°trim) | 48.423 | 0.2322 | -1.026 | 21-LT-37 | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL,1983 | RDA |
| 1982(4°trim) | 40.248 | 0.4758 | 0.09 | 25-LT-41 | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL.1983 | RDA |
| Trachurus trecae | ||||||||
| 19873–76,81–82 | 41.75 | 0.2166723 | -1.403 | 10-lf-42 | 16–26°N | OTOLITHES | FAO,1984(p53) | ROUMANIE |
| 1981 | 41.9 | 0.345 | -1.098 | 14-LT-41 | MAURITANIE | OTOLITHES | FAO,1984(p8) | RDA |
| ? | 50.2 | 0.22 | -0.59 | LF | 34.1.3 | OTOLITHES | FAO,1984(p8) | URSS |
| ? | 58.08 | 0.15 | -0.66 | LF | 34.3.1(Guinée B.) | OTOLITHES | FAO,1984(p8) | URSS |
| 1982(1°trim) | 98.805 | 0.0519 | -4.275 | 19–LT–37 | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL,1983 | RDA |
| 1982(2°trim) | 45.588 | 0.2591 | -1.161 | 24–LT–44 | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL,1983 | RDA |
| 1982(3°trim) | 43.78 | 0.3449 | 0.496 | 21-LT-45 | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL,1983 | RDA |
| 1982(4°trim) | 45.287 | 0.3168 | -1.431 | 19-LT-49 | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL,1983 | RDA |
| 1984–85 | 38.45 | 0.257 | -0.843 | 14-LF-33 | SENEGAL | EPINE DORSALE | CAMARENA,1986 | SENEGAL |
| Decapterus rhonchus | ||||||||
| ? | 48.57 | 0.16 | -0.8 | LT | 14–21°N | OTOLITHES | FAO,1979(p12) | URSS |
| ? | 45.3 | 0.303 | -0.515 | LT | ? | OTOLITHES | FAO,1979(p12) | POLOGNE |
| 1972–1974 | 55.69 | 0.136 | -1.295 | LF | SENEGAL-MAURITANIE | ECAILLES | MAXIM ET STAICU,1976 | ROUMANIE |
| ? | 44 | 0.448 | 0.114 | LF | SENEGAL | ? | FAO,1984(p12) | SENEGAL* |
| 1980 | 52.73 | 0.2713 | -0.182 | LT | MAURITANIE | OTOLITHES | HOFFMANN,1987 | RDA |
| 1981 | 58.72 | 0.1474 | -2.336 | LT | MAURITANIE | OTOLITHES | HOFFMANN,1987 | RDA |
| 1982 | 55.14 | 0.1986 | -1.18 | LT | MAURITANIE | OTOLITHES | HOFFMANN,1987 | RDA |
| 1983 | 47.8 | 0.1614 | -2.212 | LF | MAURITANIE | OTOLITHES | HOFFMANN,1987 | RDA |
| 1984 | 42.13 | 0.2426 | -1.567 | LF | MAURITANIE | OTOLITHES | HOFFMANN,1987 | RDA |
| 1985 | 51.55 | 0.1524 | -1.839 | LF | MAURITANIE | OTOLITHES | ** | RDA |
| 1982(2°trim) | 43.585 | 03461 | -0.959 | LT | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL,1983 | RDA |
| 1982(3°trim) | 48.271 | 0.2661 | -1.258 | LT | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL,1983 | RDA |
| 1982(4°trim) | 50.609 | 0.2133 | -1.592 | LT | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL,1983 | RDA |
| * Préconisé par le groupe de travail | ||||||||
| ** présenté au groupe de travail mais non publié | ||||||||
| Scomber japonicus | ||||||||
| 1972–73 | 55.357 | 0.118 | -3.178 | 29-lf-44 | 17–26°N | ECAILLES | STAICU ET MAXIM,1974 | ROUMANIE |
| ? | 44.08 | 0.326 | -0.834 | 20-LF-39 | SAHARA | ECAILLES | FAO,1979(p130) | URSS |
| ? | 48.743 | 0.201 | -2.963 | 19-LT-43 | SEN-MAURITANIE | OTOLITHES | FAO,1984(p14) | RDA |
| ? | 44.096 | 0.309 | -1.011 | LF | 34.1.3 | OTOLITHES | FAO,1984(p15) | URSS* |
| 1982(2°trim) | 44.825 | 0.339 | -1.037 | 32-lt-42 | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL,1983 | RDA |
| 1982(4°trim) | 67.515 | 0.1086 | -2.515 | 25-LT-48 | MAURITANIE | OTOLITHES | HOLZLOHNER ET AL,1983 | RDA |
| 1984–85 | 51.69 | 0.217 | -0.891 | 18–LF–41 | SENEGAL | ECAILLES | CAMARENA,1986 | SENEGAL |
* préconisé par le groupe de travail
| AIRE DE PONTE | PERIODE DE PONTE | MAXIMUM DE PONTE | PROFONDEUR (m) | TEMPERATURE OPTIMALE | TAILE DE 1° MATURITE cm | SEX-RATIO FEM/MALES | FECONDITE | SOURCES | NATIONALITE |
|---|---|---|---|---|---|---|---|---|---|
| Sardina pilchardus | |||||||||
| 34.1.3 | ttel'année | NOV-FEV | 12-13 | 1 | FAO, 1986(p82) | URSS | |||
| MAURITANIE | OCT-AVR | AVR-MAI | 15.9 | 1.15 | LAWAL ET MYLNIKOV, 1988 | MAURITANIE | |||
| Sardinella aurita | |||||||||
| 19-20°N | MARS-OCT | MAI-AOUT | 80-110 | FAO, 1979(p118) | URSS | ||||
| 13-17°N | tte l'année | MAI-AVRIL | 50-80 | 18,4-19,8 | 22 | FAO, 1979(p118) | URSS | ||
| 8-10°N | JANV-MAI | SEPT-OCT | 10-40 | 25,8-26,4 | 14 | FAO,1979(p118) | URSS | ||
| AOUT-NOV | |||||||||
| SEN-MAURIT. | tte l'année | JUIN-SEPT | 20 | 400w/gr(Sénégal) | FAO, 1979(p7) | ||||
| (déplc.du foyer | MAI Gambie | 49600-133800, | |||||||
| de reproduction) | SEPT cap Blanc | (28-40cm) | |||||||
| SENEGAL | AVR-JUIN | 18.5 | 1 | BOELY,1982 | SENEGAL | ||||
| OCT-NOV | |||||||||
| SENEGAL | MAI-JUIN | 18 | 1 | CAMARENA,1986 | SENEGAL | ||||
| NOV-DEC | |||||||||
| MAURITANIE | AVR-SEPT | 1.48 | LAWAL ET MYLNIKOV,1988 | MAURITANIE | |||||
| Sardinella maderensis | |||||||||
| SENEGAL | tte l'année | MAI-JUIL | FAO,1979(p8) | SENEGAL | |||||
| SENEGAL | JUIL-AOUT | ||||||||
| (variable) | 16.5 | BOELY,1980 | SENEGAL | ||||||
| SENEGAL | MAI-JUIL | ||||||||
| NOV-DEC | 16 | 1.2 | CAMARENA,1986 | SENEGAL | |||||
| MAURITANIE | MARS-AOUT | MAI-AOUT | 1.56 | LAWAL ET MYLNIKOV,1988 | MAURITANIE | ||||
| Engraulis encrasicolus | |||||||||
| MAURITANIE | tte l'année | AVRIL-OCTO | 16,5-24°C | males 10,4 | |||||
| femelles 10,1 | 0.56 | BA I.,1988 | MAURITANIE | ||||||
| AIRE DE PONTE | PERIODE DE PONTE | MAXIMUM DE PONTE | PROFONDEUR (m) | TEMPERATURE OPTIMALE | TAILE DE 1° MATURITE cm | SEX-RATIO FEM/MALES | FECONDITE | SOURCES | NATIONALITE |
|---|---|---|---|---|---|---|---|---|---|
| Trachurus trachurus | |||||||||
| 20-21°N(princ.) | |||||||||
| 18°30–19°30N | SEP-AVR | DEC | 110-300 | 15-16°C | 21 | 1 | 115000-127000 | FAO,1979(p9) | URSS |
| (secondaire) | |||||||||
| 14–16°N | DEC-MAI | JANV | 110-300 | 15-16°C | 16 | 115000-127000 | FAO,1979(p9) | URSS | |
| SEN-MAURIT. | OCT-MAI | DEC-FEV | 15-18°C | 19.7 | 1 | 22-346000 | FAO,1984(p6) | URSS | |
| zone dependant | (m=120000) | ||||||||
| des migrations | 18.05 | 1 | FAO,1984(p6) | ROUMANIE | |||||
| MAURITANIE | NOV-AVR | NOV-JANV | 19.3 | 1.32 | LAWAL ET MYLNIKOV,1988 | MAURITANIE | |||
| Trachurus trecae | |||||||||
| 13-19°N | FEV-MAI | BOELY ET AL,1973 | |||||||
| 21-22°N | SEP-AVR | FEV-AVR | 100-150 | 18,5-25,5°C | 31,5 | 151–772700 | FAO,1979(p10) | URSS | |
| par ponte | |||||||||
| 15-19°N | SEP-AVR | FEV-AVR | 100-150 | 18,5-25,5°C | 16,8 | FAO,1979(p10) | URSS | ||
| 11°30-14°N | SEP-AVR | FEV-AVR | 100-150 | 17,5-25,5°C | 12,5 | FAO,1979(p10) | URSS | ||
| 15-21°N | OCT-FEV | 28-30 | FAO,1984(p9) | RDA | |||||
| 15-26°N | SEPT-MARS | 24,3 | FAO,1984(p9) | URSS | |||||
| SENEGAL | FEV-AVR | 18,0 | FAO,1984(p9) | SENEGAL | |||||
| MAURITANIE | DEC-AVR | 19,56 | FAO,1984(p9) | ROUMANIE | |||||
| MAURITANIE | DEC-JUIL | JANV-AOUT | 23 | 1.1 | LAWAL ET MYLNIKOV,1988 | MAURITANIE | |||
| (tte l'année) | |||||||||
| SENEGAL | FEV-MAI | 20 | 1.4 | CAMARENA,1986 | SENEGAL | ||||
| Decapterus rhonchus | |||||||||
| SENEGAL | AVR-NOV | MAI-JUIN | 15-29°C | CONAND ET FRANQUEVILLE,1973 | |||||
| 17-22°N | JUIL-SEPT | MAXIM ET STAICU,1973 | ROUMANIE | ||||||
| 9°30-9°45N | AVR-JUIL | JUIL | 18,7°C | (2ans) | 480-990000 | FAO,1979(p127) | URSS | ||
| (m=680000) | |||||||||
| 16-21°N | JUIN-JUIL | MAI-JUIL | 18,5 | FAO,1984(p12) | URSS | ||||
| SENEGAL | AVR-NOV | MAI-JUIL | 20-21 | FAO,1984(p12) | SENEGAL | ||||
| SENEGAL | MAI-JUIN | 20 | 1.3 | CAMARENA,1986 | SENEGAL | ||||
| MAURITANIE | AVR-AOUT | AVR-AOUT | 19,4 | 1.13 | LAWAL ET MYLNIKOV,1988 | MAURITANIE | |||
| Scomber japonicus | |||||||||
| 24-25°30N, | JANV-MARS | 17,2-22,2°C | 31 | FAO,1979(p13) | URSS | ||||
| 22°30N | |||||||||
| 10-15°N | DEC-FEV | DEC | 180-200 | 17,2-22,2°C | 27.5 | FAO,1979(p13) | URSS | ||
| MAURITANIE | DEC-MARS | 31(LT) | FAO,1983(p15) | RDA | |||||
| 34.1.3 | MARS-MAI | 25.7 | FAO,1983(p15) | URSS | |||||
| 34.3.1 | DEC-MAI | 22.2 | FAO,1983(p15) | URSS | |||||
| SEN-MAURIT. | JANV-MARS | 24.36 | FAO,1983(p15) | ROUMANIE | |||||
| 1)90-250000 | FAO,1987(p19) | ? | |||||||
| 2)190-248000 | FAO,1987(p19) | ? | |||||||
| SENEGAL | 22 | *1.4 | CAMERENA,1986 | SENEGAL | |||||
| MAURITANIE | NOV-FEV | NOV-JANV | 24.6 | 1.24 | LAWAL ET MYLNIKOV,1988 | MAURITANIE | |||
| ESPECE | EQUATION | SOURCES | NATIONALITE |
|---|---|---|---|
| TRACHURUS TRACHURUS | LF=LTx0,897 | HOLZLOHNER ET KLOXIN,1985 | RDA |
| TRACHURUS TRECAE | LF=LTx0,879 | HOLZLOHNER ET KLOXIN,1985 | RDA |
| DECAPTERUS RHONCHUS | LF=LTx0,882 | HOLZLOHNER ET KLOXIN,1985 | RDA |
| SCOMBER JAPONICUS | LF=LTx0,928 | HOLZLOHNER ET KLOXIN,1985 | RDA |
| LF=0,866 LT + 1,9 | FAO,1984(p15) | URSS | |
| LF=0,8829 LT + 1,12591 | FAO,1984(p15) | RDA | |
| SARDINELLA MADERENSIS | LF=LTx0,843 | HOLZLOHNER ET KLOXIN,1985 | RDA |
| LT=1,21 LF - 0,857 | BOELY ET AL,1982 | SENEGAL | |
| SARDINELLA MADERENSIS | LF=LTx0,843 | HOLXLOHNER ET KLOXIN,1985 | RDA |
| ENGRAULIS ENCRASICOLUS | LS=0,93LF-0,41 | BA(1988) | MAURITANIE |
| LT=1,06LF+0,29 | BA(1988) | MAURITANIE | |
| (6-13 cm) | LF=1,08LS+0,04 | BA(1988) | MAURITANIE |
| LT=1,13LS+0,37 | BA(1988) | MAURITANIE | |
| LF=0,96LT-0,21 | BA(1988) | MAURITANIE | |
| LS=0,84LT-0,12 | BA(1988) | MAURITANIE |
| MORTALITE | VALEURS | METHODE | ANNEE | ZONE | SOURCE | NATIONALITE |
|---|---|---|---|---|---|---|
| Sardina pilchardus | ||||||
| M | 0.56 | PAULY | MAROC | FAO,1985(p33) | MAROC | |
| 0.65 | BEVERTON ET HOLT | MAROC | FAO,1985(p33) | MAROC | ||
| 0.6 | MAROC | FAO,1985(p33) | MAROC | |||
| 0.63 | PALOHEIMO | MAROC | FAO,1985(p33) | MAROC | ||
| 0.6 | 1972–1983 | SAHARA | FAO,1985(p92) | URSS | ||
| F | 0,17-0,37 | GULLAND | 1976–1981 | MAROC | FAO,1985(p55) | MAROC |
| 0,16-0,28 | DOUBLEDAY | 1976–1982 | MAROC | FAO,1985(p55) | MAROC | |
| Ft 0,8 | 1972-1983 | SAHARA | FAO,1985(p84) | URSS | ||
| Z | 0.8 | CHAPMAN ET ROBSON | MAROC | FAO,1985(p33) | MAROC | |
| 0.85 | COURBE DE CAPTURE | MAROC | FAO,1985(p33) | MAROC | ||
| Sardinella aurita | ||||||
| M | 0,5-0,65 | SEPT METHODES | MAURITANIE | ANON,1988(p128) | ROUMANIE | |
| MOY=0,565 | ||||||
| 0,72 | TAYLOR | SENEGAL | CAMARENA,1986(p86) | SENEGAL | ||
| 0.61-0,75 | PAULY | SENEGAL | CAMARENA,1986(p86) | SENEGAL | ||
| 0,64 | PAULY | SAH. ET MAURIT | ANON,1988(p186) | URSS | ||
| F | 0,79 | VPA | 1987 | MAURITANIE | ANON,1988(p128) | ROUMANIE |
| 0,39-0,89 | VAP | 1976–1986 | MAURITANIE | ANON,1988(p162) | ROUMANIE | |
| Sardinella maderensis | ||||||
| M | 0,54 | TAYLOR | SENEGAL | CAMARENA,1986(p86) | SENEGAL | |
| 0,53-0,66 | PAULY | SENEGAL | CAMARENA,1986(p86) | SENEGAL | ||
| Engraulis encrasicolus | ||||||
| M | 1,4-1,66 | PAULY | 1986–1988 | MAURITANIE | BA(1988) | MAURITANIE |
| F | 2,37 | BEVERTON ET HOLT | 1986–1988 | MAURITANIE | BA(1988) | MAURITANIE |
| 1986–1988 | MAURITANIE | BA(1988) | MAURITANIE | |||
| Z | 0,7-1,0 | Z=M+F | 1986–1988 | MAURITANIE | BA(1988) | MAURITANIE |
| MORTALITE | VALEURS | METHODE | ANNEE | ZONE | SOURCE | NATIONALITE |
|---|---|---|---|---|---|---|
| Trachurus trachurus | ||||||
| M | 0,5-0,77 | PAULY,LOUKASHEV, | SAHARA | FAO,1984(p13) | URSS | |
| EFANOV-RIHTER | ||||||
| 0,5 | 1985 | MAURITANIE | FAO,1987(p100) | ROUMANIE | ||
| 0,5 | 1980 | MAURITANIE | ANON.,1988(p124) | ROUMANIE | ||
| 0,5 | SAH. ET MAURIT. | ANON.,1988(p171) | URSS | |||
| 0,62 | PAULY | MAROC | FAO.,1984(p150) | MAROC | ||
| 0,65 | COURBE DE CAPTURE | 1976–1982 | MAROC | FAO, 1984 (p168) | MAROC | |
| 0,89 | CHORTES DE JONES | 1976–1982 | MAROC | FAO, 1984 (p168) | MAROC | |
| F | 0,51 | VPA | SAHARA | FAO,1984(p13) | URSS | |
| 0,62 | SAHARA | FAO,1984(p13) | ROUMANIE | |||
| 0,577 | AN.DE COHORTE | 1985 | 34.1.3,34.3.1 | FAO,1987(p100,102) | ROUMANIE | |
| 0,27-2,35(2-6 ans) | VPA | 1972–1985 | 34.1.3,34.3.1 | FAO,1987(p100) | ROUMANIE | |
| Ft | 0,427 | 1987 | MAURITANIE | ANON.,1988(p124,154) | ROUMANIE | |
| 0,16-1,31 | VPA | 1972–1987 | MAURITANIE | ANON.,1988 | ROUMANIE | |
| Ft(2-7) | 0,1-0,6 | ARMSTRONG | 1979–1981 | SAHARA | ANON.,1988(p173) | URSS |
| Fopt | 0,1-0,64(2-6ans) | CUTTY | MAURITANIE | ANON.,1988(p173) | URSS | |
| Z | 1,5-2,4 | COHORTES DE JONES | 1976–1982 | MAROC | FAO,1984(p166,167) | MAROC |
| 1,6-3,3 | COURBE DE CAPTURE | 1976–1982 | MAROC | FAO, 1984 (p166, 167) | MAROC | |
| 1, 0-1, 2 | F + M | 1972–1985 | SAHARA | FAO, 1984 (p13) | URSS | |
| Trachurus trecae | ||||||
| M | 0,4 | FAO,1983 | RDA | |||
| 0,6 | PAULY,LOUKASHEV, | SAHARA | FAO,1984(p16) | URSS | ||
| EFANOV-RIHTER | MAURITANIE | FAO,1984(p16) | ROUMANIE | |||
| 0,28 | TAYLOR | SENEGAL | CAMARENA,1986(p86) | SENEGAL | ||
| 0,54-0,67 | PAULY | SENEGAL | CAMARENA,1986(p86) | SENEGAL | ||
| 0,5 | MAURITANIE | ANON., 1988 (p125) | ROUMANIE | |||
| 0,5 | 1982–1985 | MAURITANIE | FAO, 1986 (p81, 83) | ROUMANIE | ||
| F | 0,45 | FAO, 1983 | RDA | |||
| 0,66 | VPA | 1972–1975 | SAHARA | FAO, 1984 (p17) | URSS | |
| 0,62 | 1982 | MAURITANIE | FAO, 1983 | ROUMANIE | ||
| 0,13-0,91 (2-6ans) | VPA | 1979–1987 | MAURITANIE | ANON., 1988 (p156, 125) | ROUMANIE | |
| Ft | 0,427 | 1987 | MAURITANIE | ANON., 1988 (p156) | ROUMANIE | |
| Ft | 0,11-0,86 | HOYDAL-JONES | SAH. ET MAURIT. | ANON., 1988(P175) | URSS | |
| Fort | 0,1-0,64 | THOMPSON ET BELL | SAH. ET MAURIT. | ANON., 1988(p180) | URSS | |
| Z | 1, 2 | F + M | 1972–1975 | SAHARA | FAO, 1984 (p17) | URSS |
| 0,813 | COURBE DE CAPTURE | 1982–1985 | MAURITANIE | FAO, 1986 (p81, 83) | URSS | |
| 1,34 (2-7ans) | 1982 | MAURITANIE | FAO, 1986 (p83, 84) | ANONYME | ||
| 0,835 (2-4ans) | 1982 | MAURITANIE | FAO. 1986 (p83, 84) | ANONYME | ||
| 1,964 (5-7ans) | 1982 | MAURITANIE | FAO, 1986 (p83, 84) | ANONYME | ||
| MORTALITE | VALEURS | METHODE | ANNEE | ZONE | SOURCE | NATIONALITE |
|---|---|---|---|---|---|---|
| Decapterus rhonchus | ||||||
| M | 0,45 | 5 méthodes | MAURITANIE | FAO, 1987(p110) | ROUMANIE | |
| 0,3 | PAULY | MAURITANIE | FAO, 1986(p86) | ROUMANIE | ||
| F | 0,3–0,6 | COHORTES DE JONES | 1982 | MAURITANIE | FAO,1986(p86) | ROUMANIE |
| 0,2–0,9(3–6 ans) | VPA | 1972–1985 | 34.1.3, 34.3.1 | FAO, 1987(p116) | ROUMANIE | |
| 0,494 | VPA | 1987 | MAURITANIE | ANON,1988(p126) | ROUMANIE | |
| Z | 1,77 | COURBE DE CAPTURE | 1982–1985 | SAH. et MAURIT. | FAO, 1986(p86–87) | ANONYME |
| données soviétiques | ||||||
| 1,84(5 ans et plus) | COURBE DE CAPTURE | 1982 | MAUTITANIE | FAO,1986(p86–88) | ANONYME | |
| données roumaines | ||||||
| Scomber japonicus | ||||||
| M | 0,27 | 1972 | FAO,1983 | RDA | ||
| 0,6 | PAULY, LOUKASHEV, | SAHARA | FAO,1984(p89) | URSS | ||
| TIURINE | ||||||
| 0,64 | 1982–1985 | SAHARA | ANON.,1988(p183) | URSS | ||
| 0,4 | 1972–1984 | MAURITANIE | FAO,1987(p87) | ROUMANIE | ||
| 0,23 | TAYLOR | SENEGAL | CAMARENA,1986(p86) | SENEGAL | ||
| 0,45–0,56 | PAULY | 1976–1982 | SENEGAL | CAMARENA,1986(p86) | SENEGAL | |
| 0,45 | LOUKASHEV,PAULY,GUN- | MAURITANIE | FAO,1987(p82,83) | ROUMANIE | ||
| [0,25–0,5] | DERSON,EFANOV-RIHTER | |||||
| 0,4 | MAURITANIE | ANON.,1988(p129) | ROUMANIE | |||
| F | 0,52 | FAO,1983 | RDA | |||
| 0,6 | SAHARA | FAO,1984(p9) | URSS | |||
| 0,549(2–7 ans) | VPA | 1985 | SAH. ET MAURIT. | FAO,1987(p95,97) | ROUMANIE | |
| 0,2–1,2(2–7 ans) | VPA | 1972–1983 | 34.1.3 | FAO,1987(p92) | ROUMANIE | |
| 0,4–0,7 | ANALYSE DE COHORTES | 1982–1985 | SAH. ET MAURIT. | ANON,1988(p183) | URSS | |
| 0,429 | VPA | 1987 | MAURITANIE | ANON.,1988(p129) | ROUMANIE | |
| 0,36–0,98(2–6ans) | METHODE GAMMA | 1987 | SAH. ET MAURIT. | ANON,1988(p183) | URSS | |
| Z | 1,2 | F+M | 1969 | SAHARA | FAO,1984(p9) | URSS |
| 0,62 | COURBE DE CAPTURE | 1982–1985 | MAURITANIE | FAO,1986(p85) | ANON. | |
| ESPECES | L infini cm | K | SOURCES | TEMPERATURES | MORTALITE NAT. |
|---|---|---|---|---|---|
| Sardinella aurita | 40.7 | 0.326 | PHUAM TUOC ET SZYPULA (1973) | 16 | 0.365 |
| 17 | 0.375 | ||||
| 18 | 0.385 | ||||
| Sardinella maderensis | 37.5 | 0.3 | SAMB (1986) | 16 | 0.353 |
| 17 | 0.364 | ||||
| 18 | 0.373 | ||||
| 30.34 | 0.49 | CAMARENA (1986) | 16 | 0.517 | |
| 17 | 0.532 | ||||
| 18 | 0.545 | ||||
| Sardina pilchardus | 23.08 | 0.42 | FAO (1986) Roumanie | 16 | 0.504 |
| 17 | 0.518 | ||||
| 18 | 0.533 | ||||
| Trachurus trachurus | 45.82 | 0.2006 | HOFFMAN (1987) | 16 | 0.429 |
| 17 | 0.441 | ||||
| Trachurus trecae | 41.9 | 0.345 | FAO (1984) RDA | 18 | 0.661 |
| 19 | 0.678 | ||||
| 20 | 0.694 | ||||
| Decapterus rhonchus | 51.55 | 0.152 | HOFFMANN (1987) RDA | 16 | 0.345 |
| 17 | 0.355 | ||||
| 18 | 0.365 | ||||
| 19 | 0.374 | ||||
| 42.13 | 0.242 | HOFFMANN (1987) RDA | 16 | 0.496 | |
| 17 | 0.51 | ||||
| 18 | 0.523 | ||||
| 19 | 0.536 | ||||
| Scomber japonicus | 65.51 | 0.108 | HOLZHONER ET AL. (1983) RDA | 16 | 0.258 |
| 17 | 0.266 | ||||
| 18 | 0.273 | ||||
| 19 | 0.28 |
FAO, Comité des Pêches pour l'Atlantique Centre-est4, 1984 - Rapport du Groupe de travail ad hoc sur les chinchards et les maquereaux de la zone nord du COPACE. Nouadhibou, Mauritanie, 30 janvier - 4 février 1983. COPACE/PACE Séries 83/27 : 188 p.
FAO, Comité des Pêches pour l'Atlantique Centre-est5, 1986 - Rapport de la troisième réunion du Groupe de travail ad hoc sur la sardine Sardina pilchardus (Walb.) COPACE/PACE Séries 86/39 : 157 p.
FAO, Comité des Pêches pour l'Atlantique Centre-est6, 1987 - Rapport du deuxième Groupe de travail ad hoc sur les chinchards et les maquereaux de la zone Nord du COPACE. COPACE/PACE Séries 87/46: 190 p.
Fréon P., Boély T. et Stéquert B., 1979 - Les poissons pélagiques côtiers au Sénégal. Relations taille/poids des principales espèces d'intérêt commercial. COPACE/PACE Séries 78/10 : 119–126.
Fréon P. et Stéquert B., 1979 - Note sur la présence de Sardina pilchardus (Walb) au Sénégal : Etude de la biométrie et interprétation. Cybium 3ème série, 1979(6) : 65–90.
Hoffman V., 1987(a) - Ergegnisse zu Untersuchungen an Trachurus trachurus (Linnaeus, 1758) vor Mauretanien 1980 bis 1984 Fischerei-Forschung, Rostock 25,1 : 17–22.
Hoffman V., 1987(b) - Ergebnisse zu Untersuchungen an Decapterus rhonchus (Geoffroy Saint-Hilaire, 1817) vor Mauretanien 1980 bis 1984. ibidem : 23–28.
Holzlöhner S. et Kloxin G., 1982 - On the species composition and the length - age structure of the important pelagic fisches off North West Africa, 1981. ICES/C.M. 1982/H : 55. Pelagic Fish Committee : 8 p., 2 tab., 16 fig.
Holzlöhner S. et Kloxin G., 1985 - Stock results of the most important pelagic fish species off Mauretania in 1983 and 1984. ICES/C.M. 1985/H : 72. Pelagic Fish Committee: 12 p., 5 tab., 9 fig.
Holzlöhner S., Kloxin G., Pingel C. et Hoffmann U., 1983 - On the species composition and Length/age structure of the most important fish species off Mauretania in 1982. ICES/C.M. 1983/H : 53. Pelagic Fish Committee : 44 p.
Krivospitchenko S.G., 1979 - Le maquereau Scomber japonicus devant le littoral saharien COPACE/PACE Séries 78/10 : 130–133.
Lawal H.S., et Mylnikov N., 1988 - Contribution à l'étude de la relation taille-poids de la reproduction et du sex-ratio des principales espèces pélagiques en Mauritanie : In Rapport du Groupe de travail CNROP/CRODT/ISRA sur les ressources pélagiques côtières (Mauritanie-Sénégal). Nouadhibou 8–16 juin 1988 (Miméo) : 82–97.
Maxim C. et Staicu I., 1976 - Observations sur la biologie et la dynamique du stock de saurel. (Caranx rhonchus Geof. St. Hil.) dans l' Atlantique est-central. Cercetari marine IRCM 9 : 215–226.
Moniz Carvalho M.E., 1987 - Contribution à l'étude de quelques petits pélagiques du Cap-Vert. COPACE/PACE Séries 87/46 : 119–126.
Overko S.M., 1979 - Morpho-biological characteristics of Caranx rhonchus in the eastern central Atlantic. COPACE/PACE Series 78/10 : 127–129
Pham Tuoc et Szypula J. 1973 - Biological characteristics of gilt sardine, Sardinella aurita Cuv, et Val, 1847, from northwest African coast. Act. Ichth. Pisc., 3(1) : 19–37.
Samb B., 1986 - Seasonal growth, mortality, recruitment pattern of Sardinella maderensis off Senegal. FAO/DANIDA. Denmark may, 1986.
Sedletskaia V.A., 1979 - Aires et période de reproduction de la sardinelle ronde (Sardinella aurita) et des principales autres espèces de poissons pélagiques côtiers de l' Atlantique Centre-est. COPACE/PACE Séries 78/10 : 129–134.
Staicu I. et Maxim C., 1974 - Observations sur la biologie et la dynamique du maquereau espagnol (Scomber japonicus colias Gmelin) dans l' Atlantique est-central. Cercétari marine, IRCM 7 : 113–128.
Staicu I. et Maxim C., 1984 - La pêche roumaine et l'évaluation des stocks de chinchards dans la région nord du COPACE. COPACE/PACE Séries83/27 : 52–77.
Staicu I. et Maxim C., 1986 - La pêche roumaine de la sardine Sardina pilchardus (Walbaum)) dans la région du COPACE en 1967–1983. COPACE/PACE Séries 86/39 : 97–106.
4 voir également El Ouairi M., Staicu I. et Maxim C., 1984.
5 Voir également Barkova N.A. et Domanevski L.N., Staicu I. et Maxim C., 1986.
6 Voir également Moniz Carvalho M.E., 1987.
![]()
![]()
![]()