![]()
![]()
![]()
Les avantages et inconvénients de l'élevage des tilapias en cage ont déjà été revus à plusieurs reprises (COCHE, 1975, 1978, 1979, 1982; BALARIN et HATTON, 1979; BEVERIDGE, 1987), en comparant généralement ce mode d'élevage aux méthodes traditionnelles en étang. Nous présentons ci-après les éléments se référant principalement à la production d'alevins.
Tableau 2.30. Quantité de travail (en homme - jour) nécessaire à la production de fingerlings de T. nilotica dans des piscicultures en étangs (avec ou sans hapas) et en lacs (cages et hapas) aux Philippines. (D'après ESCOVER et al, 1987).
|
|
Ecloserie en étang |
Ecloserie en cage | |
|
Caractéristiques: |
|
| |
|
|
Surface moyenne (m2) |
6707 |
486 |
|
|
Production de fingerlings (x 1000) |
477.2 |
375 |
|
|
(n/100 m2) |
71.1 |
771.7 |
|
Préparation des étangs |
111.36 |
- | |
|
Achat de matériel |
0.98 |
1.28 | |
|
Préparation et installation des filets |
3.39 |
25.79 | |
|
Achats/sélection/stockage des géniteurs |
5.57 |
4.83 | |
|
Alimentation |
40.24 |
36.98 | |
|
Fertilisation |
14.03 |
- | |
|
Nettoyage des filets/hapas |
5.67 |
29.02 | |
|
Nettoyage et réparation des digues/drainage |
24.57 |
- | |
|
Sennage |
4.72 |
- | |
|
Récolte des alevins |
66.13 |
19.71 | |
|
Triage/comptage/vente |
27.39 |
71.0 | |
|
Surveillance |
- |
114.71 | |
|
Total hommes/jours |
303.95 |
303.32 | |
|
Hommes-jour/100 m2 |
4.53 |
62.20 | |
En ce qui concerne la production d'alevins, la technique d'élevage en hapas permet d'augmenter très sensiblement la quantité de larves produites grâce à la récolte fréquente des larves au fur et à mesure de leur production. Ces récoltes, répétées et complètes, sont d'autant plus efficaces qu'elles ne nécessitent pas de vidange de l'étang, ni de pêche au filet de senne, et donc limitent les pertes de progéniture régulièrement observées lors de ces opérations. De plus, le système de cages à double filet réduit le cannibalisme exercé par les adultes, augmentant ainsi le nombre de larves produites par femelle.
La facilité de contrôle et de calibrage des alevins permet une utilisation optimale de la nourriture distribuée, d'autant plus que le taux de croissance spécifique est maximal durant les premières phases de l'élevage et que les rations doivent être adaptées à chaque classe de taille de la population. Le contrôle de l'état sanitaire des poissons est également grandement facilité.
L'importance relative des inconvénients de l'élevage en cage varie d'un endroit à l'autre. Les principaux problèmes sont liés au site d'implantation de la pisciculture, à la qualité de l'eau, à l'alimentation des alevins, à la prédation et aux maladies, aux coûts de production et enfin au vol et au vandalisme.
Qualité d'eau
Un des paramètres primordiaux concernant la qualité de l'eau est certainement la concentration en O2 dissous. L'alevinage en cage nécessite des filets à mailles fines, réduisant fortement les échanges d'eau au sein de la cage. La surveillance de la concentration en O2 dissous est donc de première importance. COCHE (1982) signale qu'à une température de 26 à 30°C, des mesures spéciales doivent être envisagées si la concentration en O2 à la surface de l'eau descend en-dessous de 3 mg/l durant plusieurs jours. Il conseille une réduction ou même un arrêt complet du nourrissage ainsi qu'une diminution de la densité de stockage et un espacement maximal des cages (à plusieurs mètres l'une de l'autre). Un entretien régulier des hapas et des cages, avec nettoyage des filets, évitera une obstruction trop importante des mailles et assurera une meilleure circulation de l'eau au sein de la cage.
Un système d'aération électrique peut éventuellement être envisagé dans les piscicultures commerciales. Dans ce cas, un système submergé semble plus efficace qu'un agitateur de surface (BEVERIDGE, 1987).
Alimentation
En ce qui concerne plus spécifiquement les poissons, mentionnons tout d'abord la haute dépendance vis-à-vis de la nourriture artificielle, surtout lorsque les densités d'élevage en alevins sont fortes. L'aliment doit être de qualité et bien équilibré d'autant plus que la croissance est très rapide durant les premiers stades et que les malformations engendrées par une carence en certains éléments (acides aminés essentiels, vitamines, ...) se manifestent de façon accélérée.
Signalons également les pertes de nourriture, soit directement à travers les mailles de la cage, soit par l'introduction d'espèces sauvages qui peuvent entrer en compétition avec les poissons en élevage. Un contrôle régulier des cages permet toutefois d'éliminer systématiquement les espèces indésirables.
Prédation et maladies
La prédation est exercée, soit par les oiseaux piscivores attirés par une densité de poissons importante, soit par les poissons voraces tels que Lates niloticus. COCHE (1982) signale également des dégâts importants provoqués par des iguanes, des tortues, des varans, des crocodiles ou des crabes. A cela, il faut ajouter les loutres capables de déchirer les filets et même les grillages de protection. Cette protection des cages est généralement réalisée à l'aide d'un filet de couverture et d'un filet submergé entourant un groupe de cages. Ces filets doivent être régulièrement contrôlés pour repérer les déchirures éventuelles.
Les conditions d'élevage en cage sont souvent considérées comme plus stressantes pour les poissons que l'élevage en étang, les rendant de la sorte moins résistants aux agents pathogènes. Peu de cas de maladies ont toutefois été recensées dans les élevages de tilapias en cage. Les quelques cas répertoriés proviennent de stress induit par des biomasses ou des salinités trop élevées (COCHE, 1982), des manipulations maladroites ou une suralimentation (PARREL et al, 1986). COCHE (1977) mentionne une diminution de la production suite aux infections mycosiques.
Coûts de production
La pisciculture en cage nécessite une série d'intrants (alimentation, matériel) qui ne peuvent être couverts que par un prix de vente du tilapia relativement élevé. Pour avoir sous-estime ce problème la culture en cage sur le barrage de Lobi au Burkina Faso a été rapidement fermée vu le déficit accumulé. L'expérience d'élevage en cage sur le fleuve Niger ne pourra se développer que si les prix du tilapia se maintiennent au niveau actuel très élevé (1.200 CFA/kg au détail).
Vol et vandalisme
Le vol constitue un problème majeur en élevage en cage. De nombreux auteurs (STREET, 1978; IDRC/SEAFDEC, 1979; COCHE, 1982; ESCOVER et CLAVERIA, 1985; BEVERIDGE, 1987) mentionnent la nécessité d'employer un gardien à temps plein et d'installer une habitation à proximité ou au milieu de l'aire de production. Ainsi, ESCOVER et CLAVERIA (1985) signalent que dans certains élevages en cage des Philippines, le vol constitue le problème principal 25% de l'ensemble des problèmes rencontrés et que la surveillance représente l'activité prépondérante (environ 50% du travail total).
Différentes stratégies de production d'alevins de T. nilotica en hapas et en cages peuvent être recommandées, selon le niveau d'intensification envisagé et la qualification technique du pisciculteur.
L'utilisation de hapas, poches fixes de petites tailles en filets moustiquaires, permet de bien contrôler et d'intensifier la reproduction et l'alevinage de T. nilotica aussi bien en étangs qu'en tanks. A une densité de 5 géniteurs/m2 avec rapport des sexes  /
/ de 3/1, on obtient des taux de recrutement très élevés, de l'ordre de 40 à 70 alevins/m2/jour, mais elle nécessite un contrôle très fréquent des hapas et un nourrissage régulier des géniteurs avec une alimentation équilibrée. Les modalités de production complète d'alevins de T. nilotica en hapas et en cages de prégrossissement sont présentées au tableau 2.31. Remarquons cependant que le niveau d'intensification préconisé nécessite un suivi continu des poissons en croissance (ration et fréquence d'alimentation élevées, état de santé, etc.).
de 3/1, on obtient des taux de recrutement très élevés, de l'ordre de 40 à 70 alevins/m2/jour, mais elle nécessite un contrôle très fréquent des hapas et un nourrissage régulier des géniteurs avec une alimentation équilibrée. Les modalités de production complète d'alevins de T. nilotica en hapas et en cages de prégrossissement sont présentées au tableau 2.31. Remarquons cependant que le niveau d'intensification préconisé nécessite un suivi continu des poissons en croissance (ration et fréquence d'alimentation élevées, état de santé, etc.).
Une autre méthode relativement simple et procurant des résultats satisfaisants consiste à réaliser la production des larves en étang et à les y maintenir jusqu'à ce qu'elles atteignent un poids d'environ 1g. Cette étape peut elle-même être subdivisée en 2 phases, la reproduction s'effectuant dans des petits étangs de ponte reliés via une buse de transfert à des étangs de premier alevinage (CAMPBELL, 1985). Les alevins sont ensuite déversés en cages de prégrossissement phase 1 jusqu'à 10 g, puis en cages de prégrossissement phase 2 jusqu'à 30g. La récolte des alevins de 4g, par sennage de l'étang, permet de réduire le prégrossissement en cage à une seule phase, jusqu'à ce que les poissons atteignent un poids de 20-30g, mais les taux de production en alevins sont généralement moins élevés.
Enfin un autre système de production d'alevins qui s'effectue complètement en cages flottantes de 16m2 avec filets à mailles fines consiste à y disposer quatre petites cages à géniteurs (10 ind/m2 avec rapport des sexes / = 2) avec 5 seaux à reproduction ce qui donne avec une alimentation intensive des productions de l'ordre de 400 alevins/m2/mois.
Tableau 2.31. Modalité de production d'alevins de T. nilotica en hapas et en cages de prégrossissement.
|
Caractéristiques de l'élevage
|
Phases de production |
|||
|
Reproduction |
Prégrossissement 1 |
Prégrossissement 2 |
||
|
Cages: |
||||
|
|
- dimension (m3) |
1.5-9 |
0.5-1 |
1-5 |
|
|
- mailles filet (mm) |
1-3 |
4 |
4-8 |
|
Mise en charge |
||||
|
|
- densité (ind/m3) |
5 |
3000 |
800-1300 |
|
|
- rapport des sexes ( |
3/1 |
- |
- |
|
|
- poids moyen géniteurs (g) |
100-200 |
- |
- |
|
Récolte: |
||||
|
|
- fréquence de récolte (j) |
10-14 |
- |
- |
|
|
- durée d'élevage (j) |
- |
20-50 |
30-40 |
|
|
- poids moyen alevins (g) |
<0.5 |
1-4 |
10-30 |
|
Alimentation: |
||||
|
|
- qualité (% Protéines brutes) |
20-30 |
35 |
25-30 |
|
|
- ration (% biomasse) |
1.5-3 |
15-30 |
6-10 |
|
|
- fréquence (n/j) |
1 |
4-6 |
3-4 |
Tableau 2.32. Caractéristiques d'une unité d'élevage intensif idéale. (D'après BALARIN et HALLER, 1982).
|
1. Intérieur lisse: prévention contre les dommages causés aux poissons (aspérités) et facilité de nettoyage, réduction de l'incrustation d'algues, bactéries,... |
2.3.1. Caractéristiques et construction des tanks, "raceways" et arènes
2.3.2. Gestion des stocks de géniteurs et d'alevins
2.3.3. Alimentation des géniteurs et des alevins
2.3.4. Bilan économique de la production d'alevins en tanks
2.3.5. Avantages et inconvénients de l'élevage en tanks, raceways et arènes
2.3.6. Recommandations pour la production d'alevins en tanks, "raceways" et arènes
Le développement de l'élevage intensif de T. nilotica en tanks (= bassins) n'a réellement pris son essor qu'à la fin des années 1970. BALARIN et HATTON (1979) signalent d'ailleurs qu'à l'exception des rapports de UCHIDA et KING (1962) et de HIDA et al (1962) sur l'utilisation respectivement d'auges et de "raceways" destinés à l'élevage de T. mossambica comme poisson-appât à Hawaï, peu d'essais avaient jusque là utilisé les tanks comme système d'élevage des tilapias. Depuis, les élevages en tanks se sont multipliés, tant au niveau expérimental que commercial (BALARIN et HALLER, 1982; MELARD, 1986). Ces structures d'élevage ne sont toutefois utilisables qu'en pisciculture commerciale ou industrielle, à grande échelle et donc généralement hors de portée des villageois.
Ce mode d'élevage présente l'avantage de ne nécessiter que peu d'espace, comparé à l'élevage en étang. Il permet un contrôle efficace des conditions environnementales et une gestion très précise de la production. La possibilité de produire une quantité maximale d'individus dans un volume d'eau minimal est toutefois étroitement dépendante de paramètres tels que l'alimentation (qualitative et quantitative), l'oxygénation (par aération ou renouvellement rapide du volume d'eau) et la formation du personnel employé.
BALARIN et HALLER (1982) classifient les tanks et raceways de la façon suivante:
- tanks: en béton ou en polyester de forme variable (circulaire, ovale, carrée,...), utilisés pour la production d'alevins, pour le traitement d'inversion du sexe aux androgènes (BALARIN et HATTON, 1979; GUERRERO, 1976), mais principalement destinés au grossissement et à la production de poissons de taille marchande. (BALARIN et HATTON, 1979; LAUENSTEIN. 1978; NASH et MAYO, 1979; MELARD. 1986).- raceways: en béton de forme généralement rectangulaire, principalement utilisés pour l'élevage des alevins, soit jusqu'à une taille commercialisable comme poissons-appâts (UCHIDA et KING, 1962), soit comme stabulation avant le déversement en étangs (RAY, 1978), en tanks de grossissement (LAUENSTEIN, 1978; BALARIN et HALLER, 1979) ou en cages (KONIKOFF, 1975).
- les arènes, développées par HALLER et PARKER (1981): plateformes circulaires servant d'aire de reproduction et occupées en continu par les mâles qui y construisent des nids et ponctuellement par les femelles qui viennent y pondre.
2.3.1.1. Modèles de tanks et raceways.
2.3.1.2. Arènes de reproduction
2.3.1.3. Dispositif et taux d'écoulement
Comparés aux étangs et éventuellement aux raceways, les tanks sont généralement de petite taille et construits en béton, bois, fibre de verre ou métal. Quel que soit le matériau utilisé, leur surface intérieure doit être aussi lisse que possible, facilitant de la sorte les opérations de nettoyage tout en empêchant les poissons de se blesser aux aspérités des parois. Les autres caractéristiques d'un tank d'élevage idéal sont présentées au tableau 2.32. En général, l'alimentation en eau des tanks, raceways et arènes est effectuée par pompage, ce qui implique, à la différence des étangs alimentés en eau par gravité, un coût énergétique récurrent qu'il faut bien évaluer. Pour la vidange, tanks, raceways et arènes doivent être positionnés pour permettre l'évacuation facile de l'eau par inclinaison ou abaissement du tuyau d'évacuation.
Les différents modèles de tanks existant sur le marché ont été inventoriés par BALARIN et HALLER (1982). Le tableau 2.33 donne les caractéristiques des modèles (planche 2.3) pouvant être recommandés pour la production et le grossissement d'alevins de tilapias.
Tableau 2.33. Caractéristiques de quelques modèles de tanks adaptés à l'élevage des larves et des alevins et du système de vidange fréquement utilisé (Modifié d'après BALARIN et HALLER, 1982).
|
Caractéristiques |
Modèles de structure |
Système de vidange | ||
|
Représentation |
1. circulaire ou carré |
2. rectangulaire |
3. raceways |
|
|
|
|
|
|
|
|
Efficacité de nettoyage |
Très bonne, système autonettoyant |
Faible, création de "zones mortes" |
Faible à bonne, nécessite un courant uniforme |
Faible à modérée; bonne quand le système est baissé manuellement |
|
Distribution de l'aliment |
très bonne |
moyenne |
bonne, mais inférieure à 1 |
- |
|
Perte d'aliment |
moyenne à élevée |
faible |
faible |
variable |
|
Distribution des poissons |
très bonne |
faible, effet d'entassement dans les coins chez les poissons de plus grande taille. |
bonne, mais souvent même réaction qu'en 2 |
- |
|
Récolte des alevins |
moyenne |
simple |
très simple |
très bonne |
|
Coût de construction |
modéré, fonction du matériel |
faible, 12% en moins que 1. |
faible, construction simple. |
- |
Les avantages du tank circulaire par rapport au tank rectangulaire ont été largement décrits par BALARIN et HALLER (1982). Rappelons que les principaux inconvénients du tank rectangulaire sont:
- présence de zones mortes,- réduction de la vitesse du flux d'eau par rapport au tank circulaire, conduisant à des déficits localisés en oxygène ainsi qu'à l'accumulation de déchets et de nourriture non ingérés, source d'agents infectieux, de stress et finalement de mort du poisson.
Ces auteurs signalent toutefois que le tank rectangulaire peut être utilisé pour l'élevage des alevins, car ceux-ci ne présentent généralement pas le comportement perturbé des poissons sauvages lorsqu'ils sont placés dans ce type de tank (entassement dans les coins du tank).
BURROWS et CHENOWETH (1955), comparant les modes d'écoulement dans les tanks circulaires, rectangulaires et ovales, ont montré que la circulation de l'eau dans les raceways était fort différente et dépendante des dispositifs d'amenée et d'évacuation d'eau. Ceux-ci doivent en effet couvrir l'entièreté de la largeur du raceway. Un rapport longueur-largeur-profondeur du raceway de 30: 3: 1 est requis pour l'évacuation efficace des déchets (WESTERS et PRATT, 1977).
HALLER et PARKER (1981) ont conçu et testé, dans une ferme piscicole du Kenya (Baobab Farm), un système de tank spécialement mis au point pour la reproduction des tilapias. Ce système se base sur la particularité des comportements de reproduction et d'incubation des tilapias incubateurs buccaux, tels qu'ils ont pu être observés en milieu naturel et décrit à plusieurs reprises (FRYER et ILES, 1972; RUWET et al, 1975). Il est constitué:
- d'une aire de reproduction appelée "arène", où les mâles délimitent chacun un territoire sur lequel ils creusent leur nid en forme de dépression et où ils invitent à la reproduction les femelles de passage,- d'une aire d'incubation et de gardiennage, en dehors de l'aire de reproduction, où la femelle peut incuber ses oeufs et veiller à sa progéniture durant les jours qui suivent l'éclosion, et
- d'une aire d'alevinage, de faible profondeur vers laquelle les alevins se dirigent spontanément à la fin de leur phase de gardiennage.
Ce système permet de la sorte d'éviter ou de réduire l'aggressivité et les attaques observées entre adultes ainsi que le cannibalisme effectué à l'égard des alevins. La figure 2.8 présente de façon détaillée le dispositif de HALLER et PARKER. La zone A constitue l'arène de reproduction (d = 10 m) et est séparée de l'aire de gardiennage B (largeur = 3.7 m) par une série de portes grillagées conçues de manière à laisser le libre passage aux femelles et à retenir les mâles choisis de taille supérieure aux femelles. La séparation entre la zone B et la zone C (largeur = 0.3 m) vers où se dirigent les alevins est constituée d'un ensemble de créneaux en forme de V pointés vers C (système de passe à poissons unidirectionnelle). Les passages ont une ouverture maximale de 0.5 cm et ne peuvent donc être utilisés que par les jeunes alevins. De plus, la profondeur de l'eau n'y excède pas 1 cm. Un abaissement du niveau d'eau de 1 cm isole les alevins se trouvant dans la zone C du reste du système et facilite leur récupération. Cette installation est mise en charge avec 280 femelles et 40 mâles (rapport des sexes /: 7/1). La vidange bi-hebdomadaire de l'aire d'alevinage (C) permet la récolte de jeunes alevins de même âge et de taille uniforme. La production d'une arène de ce type est de 200 alevins/m2/an (BALARIN et HALLER, 1983) soit 100 alevins/femelle/an ce qui est faible. Il est vrai qu'un certain nombre d'alevins ne se dirigent pas vers l'aire d'alevinage et deviennent alors progressivement cannibales vis-à-vis des alevins fraîchement éclos. Leur nombre peut être réduit en plaçant des nasses dont l'entrée est trop faible pour les adultes. Des copies réduites de ces systèmes sont parfois construites avec des moyens financiers limités. Si l'arène de reproduction n'a qu'un diamètre de 1 à 2 m, la production d'alevins ne peut justifier le coût d'une telle installation. De toute façon, malgré son intérêt pratique, ce système ne semble pas se généraliser en Afrique vu le coût des infrastructures en béton et des installations de pompage.
L'évacuation des déchets et les besoins en O2 sont considérés comme les paramètres principaux fixant le taux d'écoulement dans les tanks. Il est évidemment nécessaire d'établir un équilibre entre le niveau de nettoyage du tank et les effets du courant sur la croissance et la consommation d'O2. Ainsi, dans un circuit ouvert, un débit de 0.5 à 11/kg de poisson/min maintient un niveau d'O2 et une efficacité de nettoyage satisfaisants. BALARIN et HALLER (1982) ont établi les limites du flux d'eau pour le tilapia de la façon suivante:
- vitesse minimale du courant: 7.5-10 cm/sec, correspondant à une vitesse permettant la sédimentation des particules faecales les plus grosses.- vitesse maximale du courant: 20-30 cm/sec, correspondant à une vitesse au-dessus de laquelle les poissons sont forcés de nager.
Dans un tank circulaire, la vitesse du courant est fortement influencée par l'angle du flux d'entrée et par la profondeur du tank (LARMOYEUX et al, 1973, in BALARIN et HALLER, 1982). L'évacuation des déchets sera donc d'autant plus efficace que l'arrivée d'eau se fait de façon tangentielle.
En raceways, une pratique courante dans l'élevage des tilapias est d'ajuster la profondeur de manière à permettre un renouvellement d'eau minimum 4 fois/h, selon l'équation suivante (BALARIN et HALLER, 1982):
d = F/(A×R)
où d = profondeur utile (en m),
F = débit (m3/h),
A = surface du raceway (m2)
R = taux de renouvellement (n/h) minimum = 4.
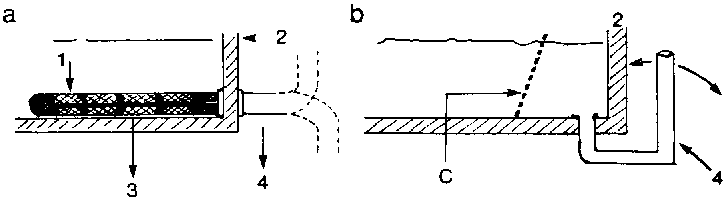
Les grands volumes d'eau généralement utilisés en circuit ouvert nécessitent un dispositif d'évacuation adéquat, le débit de sortie devant impérativement égaler le débit d'entrée afin d'éviter le débordement du basssin et la fuite des poissons. Le diamètre de la canalisation doit donc être suffisamment large pour faciliter l'écoulement du flux maximal requis lors des charges optimales du tank. Lors de l'élevage des jeunes larves, le tuyau d'évacuation doit être muni d'un filet à fines mailles empêchant le passage des poissons.
Des systèmes tels que ceux décrits à la figure 2.9 évitent une obstruction trop rapide du tuyau de vidange. Un trop-plein de sécurité doit également être prévu légèrement au-dessus du niveau normal de l'eau du tank. La taille des mailles selon l'âge, la taille et le poids des alevins de T. nilotica est mentionnée au tableau 2.34.
Tableau 2.34. Dimensions des mailles du filtre entourant le système de vidange, selon l'âge et la taille de Tilapia nilotica (D'après GUERRERO, 1985).
|
Taille de la maille (mm)
|
Long. totale (cm)
|
Caractéristiques des poissons |
|
|
Poids (g) |
Age (semaines) |
||
|
3 |
1.6 |
0.06-0.1 |
2 |
|
4 |
2.1 |
0.2-0.4 |
3 |
|
6 |
2.6 |
0.5-1.0 |
4 |
|
9 |
4.7 |
1.1-1.5 |
5 |
|
11 |
5.7 |
2.0-3.5 |
6 |
2.3.2.1. Densité de stockage et rapport des sexes
2.3.2.2. Récolte du frai
2.3.2.3. Croissance des alevins en tanks
La production d'alevins en tank est influencée par de nombreux paramètres, tels que densité et rapport des sexes des géniteurs, conditions physico-chimiques de stockage, techniques de gestion des tanks et des poissons, qualité de la nourriture,... Toutefois, comparativement à d'autres systèmes d'élevage (étangs, cages), l'élevage en tank permet un contrôle plus précis de ces différents paramètres.
Comparant des densités de stockage de 4, 7 et 10 géniteurs/m2. BAUTISTA (1987) signale que la production de larves par géniteurs est la plus élevée avec une densité de 4 ind/m2 (figure 2.10). Par unité de surface, le recrutement en alevins est toutefois supérieur avec une densité de 10 ind/m2 Une mise en charge trop élevée augmente l'aggressivité entre mâles territoriaux, réduisant de la sorte le temps consacré à la parade nuptiale (BALARIN et HALLER, 1982). A hautes densités, certains mâles ne peuvent prendre part à la reproduction par manque de territoire et donc d'aire de ponte. Le pourcentage de femelles en reproduction augmente également lorsque leur nombre par aire de ponte diminue (LOVSHIN, 1982). Des expériences menées à hautes densités de stockage en géniteurs (MELARD, 1986) indiquent que des reproductions sont observées jusqu'à 125 ind/m2 mais que la quantité d'alevins produits est faible (200 à 250 alevins par tanks de 4 m2 pendant une période de 30 jours, soit 2 alevins/m2/jour). A partir de 250 ind/m2. plus aucune reproduction n'est observée (tableau 2.35) D'après MELARD (1986), ce faible recrutement en alevins observé à haute densité ne peut être attribué uniquement au cannibalisme, mais également à un phénomène d'inhibition de la reproduction (atrésie folliculaire) avec détournement de l'énergie normalement utilisée pour la formation des oeufs vers la croissance somatique.
Lorsque le rapport des sexes augmente au-delà de 3/1, un nombre croissant de femelles ne participent pas à la reproduction et, soit exercent une prédation sur les jeunes alevins, soit perturbent la ponte des autres femelles en tentant d'ingérer les oeufs à mesure qu'ils sont pondus.
MELARD et PHILIPPART (1981) rapportent par contre que le meilleur rapport femelle/mâle est de 5/1, aussi bien en tank qu'en étang ou en cage. D'autre part, HALLER et PARKER (1981) utilisant le système des arènes préconisent un rapport femelle T. nilotica/mâle T. aurea de 7/1, sans observer pour autant les problèmes décrits précédemment. BALARIN et HALLER (1982) signalent qu'avec un rapport / inférieur à 3/1, le nombre de femelles est insuffisant pour permettre une reproduction continue à partir des mâles disponibles, alors que GUERRERO et GUERRERO (1984) recommandent une densité de 3 géniteurs/m2 avec un rapport / de 2/1 (tableau 2.36).
La taille des femelles utilisées est généralement faible, inférieure à 200 g (BALARIN et HALLER, 1982; GUERRERO et GUERRERO, 1985). MELARD et PHILIPPART (1981), de même que COCHE (1982), recommandent par contre des femelles de 150 à 400 g. De ces informations parfois contradictoires il est difficile de recommander une pratique plutôt qu'une autre. Toutefois, des informations recueillies auprès de pisciculteurs producteurs (non chercheurs), il apparaît qu'une densité de l'ordre de 4 ind/m2 avec un rapport des sexes / de 3/1 et en utilisant de préférence des femelles de poids supérieur à 200 g (à condition d'avoir des mâles un peu plus gros) donne de bons résultats.
Tableau 2.35. Effet de la densité de population sur la territorialité, la production d'alevins et la croissance de Tilapia nilotica (T° = 23°C, tank de 4 m2/l m3) (d'après MELARD, 1986).
|
Densité (ind/m2) |
50 |
75 |
125 |
250 |
500 |
750 |
|
Pmi(g) |
9.9 |
9.6 |
9.1 |
9.6 |
9.2 |
9.4 |
|
Pmf |
83.2 |
83.8 |
83.2 |
80.2 |
68.3 |
62.7 |
|
Pmf |
44.1 |
44.6 |
46.4 |
49.3 |
44.5 |
41.3 |
|
Rapport des sexes ( |
1.04 |
1.03 |
1.12 |
1.02 |
1.10 |
1.11 |
|
% mâles territoriaux |
7.8 |
3.2 |
3.4 |
0.3 |
0.1 |
0.0 |
|
Production d'alevins (n/m2/jour) |
4.0 |
3.4 |
2.0 |
0.0 |
0.0 |
0.0 |
Figure 2.11. Variation de la ration journalière (% biomasse/j) en fonction du poids des alevins de tilapias.
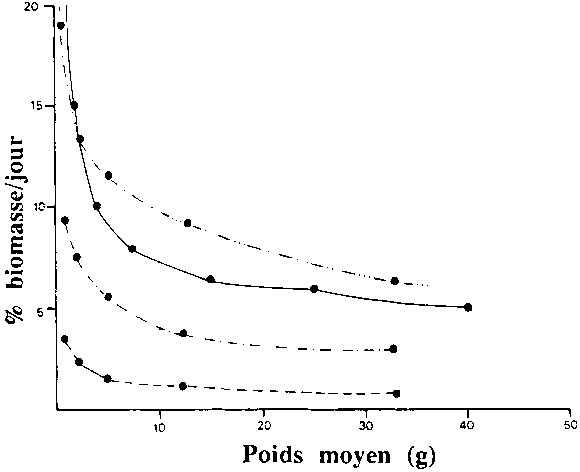
- calculé d'après SARIG et MAREK, 1974 - aliment: 25% protéines, t° 20-28°C
- ration maximale d'après MELARD, 1986 - aliment: 46-50% PB, t° 25-27°C
- ration de maintenance d'après MELARD, 1986 - aliment: 46-50% PB, t° 25-27°C
- ration optimale d'après MELARD, 1986 - aliment: 46-50% PB, t° 25-27°C
Tableau 2.36. Production de larves de Tilapia nilotica (ou d'espèces proches) en tanks en fonction de la densité des géniteurs, du rapport des sexes et de la fréquence de récolte.
|
Espèces
|
Densité de géniteurs (n/m3)
|
Rapport
|
Pmi
|
Période de production (jours)
|
Fréq. de récolte
|
Production |
Commentaires
|
Références
|
|
|
(n/m2/j) |
(n/kg |
||||||||
|
T. nilotica |
4.5 |
5:1 |
208 |
50 |
1/j |
36 |
46.5 |
Bassin de 4m2/1m3 circuit fermé, eaux réchauffées par centrale nucléaire |
MELARD et PHILIPPART 1981 |
|
8.3 |
10:1 |
98 |
57 |
1/J |
25 |
34 |
|
|
|
|
3 |
2:1 |
95-120 |
20.7 |
4-6/j |
16.2 |
62 |
Bassin de 120m2 pic de reprod. |
GUERRERO & GUERRERO 1985 |
|
|
3 |
2:1 |
95-120 |
60.7 |
4-6/j |
9 |
34.9 |
Cycle de reprod. Complet |
|
|
|
8.4 |
3.75:1 |
300 |
- |
1/15j |
67-90 |
38.5-44 |
Bassin de 2.25m2/1.35m3 eaux réchauffées |
McANDREW (pers. com., in BALARIN et HALLER, 1982) |
|
|
4.4-6.5 |
5:1 |
200-400 |
- |
1/1j |
23-46 |
17-34 |
Bassin de 55m2 eau légèrement saumâtre (4 à 8‰) |
CAMPBELL (pers. com., in COCHE 1982) |
|
|
T aurea × T. nilotica |
20 |
8-10:1 |
50-80 |
- |
1/1 |
- |
42.7-85.4 |
Bassin de 5001 avec graviers. |
MIRES 1977 |
|
2 |
7:1 |
80 |
- |
1/3.5j |
6-7 |
43-50 |
Bassin de 154m2 avec arènes de reproduction. |
HALLER et PARKER. 1981 |
|
|
T. aurea |
1.6 |
3:1 |
127-284 |
90 |
1/7j |
23 |
67-150 |
Bassin de 7.3m2 |
SNOW et al, 1983 |
|
|
|
|
|
|
|
|
Capture à l'épuis. et à la senne (A) |
|
|
|
1.6 |
3:1 |
127-284 |
90 |
1/7j |
17.6 |
52-117 |
idem que A," + vidange ttes les 3 semaines |
|
|
|
1.6 |
3:1 |
127-284 |
90 |
1/10-12j |
24 |
71-159 |
Sennage tous les 10-12 jours et récolte des oeufs dans la bouche des femelles. |
|
|
|
1.6 |
3:1 |
127-284 |
90 |
1/90j |
0.7 |
2-4.4 |
Aucune récolte intermédiaire |
|
|
|
10 |
3:1 |
13-18 cm |
- |
1/1j |
7 |
- |
Bassin de 18 m2 |
SHELTON et al, 1978 |
|
Différentes méthodes de gestion des tanks de reproduction ont été expérimentées afin d'augmenter la production des larves par femelle et par unité de surface.
La récolte régulière des larves est habituellement effectuée à l'aide d'épuisettes à fines mailles. La capture des géniteurs peut éventuellement être réalisée par sennage des tanks, lorsque ceux-ci sont de grandes dimensions. Cette capture permet d'examiner l'état des géniteurs et de prélever les oeufs en incubation dans la bouche des femelles.
Une vidange régulière du tank (à intervalle de 3 semaines à 1 mois) peut également être envisagée afin de maintenir une qualité d'eau satisfaisante, du moins lorsqu'aucun système de renouvellement d'eau n'est prévu (SNOW et al, 1983; GUERRERO et GUERRERO, 1985).
La première récolte des larves, après mise en charge des tanks avec des géniteurs à maturité, est généralement effectuée entre le 11ème et le 34ème jour (MELARD et PHILIPPART, 1981; GUERRERO et GUERRERO, 1985). La fréquence de récolte des larves peut varier fortement selon les auteurs. Ainsi, CAMPBELL (com. pers. in COCHE, 1982) et GUERRERO et GUERRERO (1985) recommandent une récolte journalière des larves de tilapias, ou même plusieurs fois par jour, alors que SNOW et al (1983) n'effectuent la récolte que tous les 10 à 12 jours ou même toutes les trois semaines (Tableau 2.36). La récolte journalière, facilement réalisable en tank, présente plusieurs avantages:
- les jeunes alevins au stade de la nage libre se déplacent en "nuage",
- la prédation des jeunes par les adultes est réduite au minimum,
- les soins à apporter aux oeufs et aux larves vésiculées sont supprimés.
Le principal inconvénient est le coût en personnel, quoique cela ne représente pas réellement un facteur limitant dans les pays en voie de développement Notons que la récolte des jeunes en tanks ne présente pas le problème des pertes importantes en alevins (30%) signalées par ROTHBARD et al (1983) lors de la vidange régulière des étangs de reproduction. La récolte des oeufs, directement dans la bouche des femelles de T. nilotica et leur incubation artificielle a déjà été rapportée par NUSSBAUM et CHERVINSKY (1968). SNOW et al (1983), travaillant avec T. aurea, considèrent que le temps investi à cette opération n'est pas justifié par une augmentation significative du nombre d'alevins produits (Tableau 2.36). HOERTGEN-SCHWARK et al (1987) ont toutefois montré que l'intervalle de temps entre 2 pontes successives pouvait être réduit de 7 jours lors d'une incubation artificielle des oeufs.
Le remplacement régulier des géniteurs peut également améliorer les résultats de production. LOVSHIN (1980) a en effet montré que la production de larves diminuait considérablement après le troisième cycle de ponte, chaque cycle durant environ 2.5 mois. Parallèlement, GUERRERO et GUERRERO (1985) ont démontré que, pour une période de reproduction s'étalant sur 50 à 72 jours, un pic de pontes était observé après environ 21 jours (16.2 alevins/m2/jour) alors que la production moyenne calculée sur l'ensemble de la période de reproduction ne dépassait pas 9 alevins/m2/jour (Tableau 2.36). D'après leurs résultats, il semble donc plus intéressant de séparer les couples de T. nilotica après le pic de production d'alevins plutôt que de les maintenir durant des temps plus longs. Les tanks peuvent à nouveau être mis en charge avec de nouveaux géniteurs.
Les productions d'alevins les plus élevées (McANDREW, 1988) sont probablement obtenues dans le système combiné de grands tanks divisés en différents compartiments par des hapas. Ce système permet un contrôle facile des géniteurs qui sont suivis au niveau individuel. Cette méthode permet une récupération manuelle totale des alevins de chaque femelle qui est alors reconditionnée dans un tank monosexe femelle par alimentation intensive. Un marquage individuel des femelles permet de sélectionner les plus fertiles et d'éliminer les individus stériles. La production d'alevins selon cette technique varie entre 2.200 et 3.000 ind/m2/mois (McANDREW, 1988).
Les larves, régulièrement prélevées à partir des tanks de reproduction, sont déversées en tanks de prégrossissement (planche 2.3) jusqu'à une taille d'environ 30 g. Cette phase de prégrossissement est souvent réalisée dans une série de tanks successifs, avec recalibrage régulier des alevins et diminution progressive de la densité de stockage. Lorsqu'ils atteignent un poids de quelques g, les jeunes alevins peuvent également être déversés en cage de grossissement (COCHE, 1982).
BALARIN et HALLER (1979) recommandent le processus de mise en charge suivant: les alevins de 1 à 5 g sont stockés à une densité de 1600 ind/m3, densité progressivement réduite par triage sélectif pour atteindre 1000 poissons de 10 à 20 g/m3 après 30 à 40 jours. Une seconde étape de croissance de 30 jours permet l'obtention de "fingerlings" de 25 à 50 g, avec une biomasse d'environ 35 kg/m3 Par la technique du triage sélectif, seulement 50% de la population initiale, à savoir les poissons à croissance rapide, sont placés en grossissement. Parmi ceux-ci, on retrouve généralement 70 à 80% d'individus mâles. Les autres peuvent être vendus comme poissons-amorces, pour la mise en charge d'étang ou comme source de protéines animales dans l'alimentation pour animaux (poissons, porcs) et humains (fretin) (BALARIN et HALLER, 1982; 1983).
Le triage sélectif peut être réalisé dans des raceways rectangulaires, le système de triage étant constitué d'un grillage en forme de sac dont l'entrée est de dimension égale à la largeur du raceway et qui peut ainsi être déplacé tout au long du tank en ne retenant que les individus de grande taille. D'autres systèmes ont également été décrits par BALARIN et HALLER (1982) et ARMITAGE et al (1987). L'espacement des grilles est imposé par l'épaisseur du poisson (Tableau 2.37).
Tableau 2.37. Epaisseur du corps de T. nilotica élevé en tank et dimensions recommandées pour l'écartement des grilles de triage
|
Poids moyen (g)
|
Epaisseur de T. nilotica (cm) |
Espace des grilles (cm)
|
|
|
a) |
b) |
||
|
>5 |
0.99 |
0.97 |
1.00 |
|
10 |
1.23 |
1.19 |
1.25 |
|
25 |
1.65 |
1.40 |
1.70 |
a) d'après BALARIN et HALLER, 1982;
b) d'après PRUGININ et SHELL, 1962.
Des densités de démarrage de 2000 à 4000 larves/m2 ont été rapportées par CAMPBELL (com. pers., in COCHE, 1982), la hauteur d'eau dans les tanks étant progressivement élevée au fur et à mesure de la croissance des alevins. Plus de 30000 alevins de 2 à 3 g peuvent ainsi être produits dans un tank de 4x4 m, avec un renouvellement d'eau important. La capacité de charge optimale d'un tank d'élevage est en effet fortement influencée par le taux de renouvellement d'eau dans le tank. Ainsi, MELARD et PHILIPPART (1981) proposent différentes charges optimales pour la croissance de T. nilotica, en fonction du poids moyen des alevins et du taux de renouvellement d'eau (Tableau 2.38).
Tableau 2.38. Biomasse en kg et nombre par m3 et charges optimales calculées pour différentes catégories de T. nilotica élevés en tank de 1 m3. à une température de 30-32 °C (d'après MELARD et PHILIPPART, 1981) (): n/m3
|
Poids moyen (g)
|
Charge (kg/l/min)
|
Biomasse (kg/m3) pour un renouvellement de l'eau de: |
|||
|
1x/h |
2x/h |
3x/h |
4x/h |
||
|
1 |
0.07 |
1.17(1170) |
2.34 |
3.51 |
4.68 |
|
10 |
0.29 |
4.83 (483) |
9.66 |
14.49 |
19.32 |
|
50 |
0.77 |
12.83 (257) |
25.66 |
38.49 |
51.32 |
Les taux de production moyen d'alevins de T. nilotica obtenus par différents auteurs travaillant en élevage intensif en tank sont présentés au tableau 2.39. Nous pouvons ainsi observer des mises en charge particulièrement élevées (12350 alevins de 1-5 g/m3) lors de l'apport d'une alimentation artificielle riche en protéines (45%) et avec un taux de renouvellement d'eau important (2.5 fois/h). Toutefois, au fur et à mesure de l'accroissement en poids les mises en charge diminuent nettement (1000 à 2240 alevins de 10-20 g/m3).
Tableau 2.39. Prégrossissement des alevins de T. nilotica en élevage intensif en tank.
|
Mise en charge |
Durée d'élevage |
Vidange |
Production |
Aliment |
Système de production
|
Références
|
|||
|
Pmi (g) |
n/m3 |
Bi (kg) |
(j) |
Pmf (m) |
Bf (kg) |
(kg/m3/mois) |
type-ration |
||
|
1-5 |
1650 |
5.0 |
40-50 |
10-20 |
22 |
9.5 |
granulés |
Raceway 15m2/6m3 renouvel. 3-4/h |
BALARIN et HALLER, 1979 |
|
10-20 |
1000 |
14.0 |
40-50 |
25-50 |
35.6 |
11.7 |
granulés |
|
|
|
1-5 |
12350 |
30.9 |
82 |
5-10 |
92.6 |
22.5 |
granulés |
Bassin 4m2/1m3 renouv. 2.5/h |
MELARD et PHILIPPART, 1981 |
|
3.1 |
1000 |
3.1 |
31 |
17.7 |
17.6 |
14.0 |
granulés |
renouvel. 0.9/h |
|
|
9.1 |
500 |
4.6 |
45 |
63.9 |
31.9 |
39.5 |
gr. 45% PB |
renouv. 1.3/h |
MELARD, 1986 |
|
10-20 |
2240 |
- |
- |
20-50 |
- |
10.5 |
gr. 45% PB |
Bassin 4m2/1m3 renouv. 2.3/h |
|
Pmi, f = Poids moyen initial et final
Bi, f = Biomasse initiale et finale
![]()
![]()
![]()



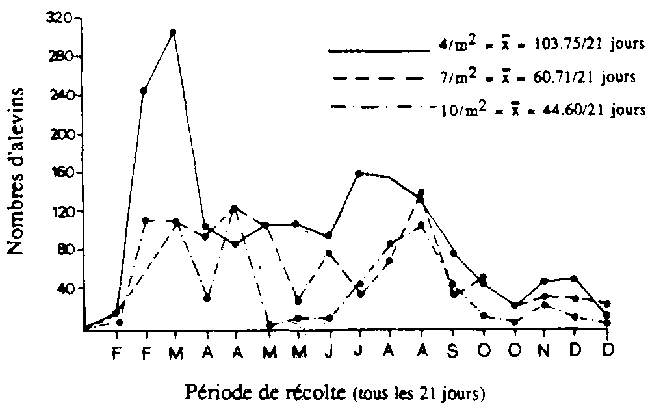 ) de Tilapia nilotica (d'après BAUTISTA, 1987). x = nombre moyen d'alevins produits par femelle durant une période de 21 jours
) de Tilapia nilotica (d'après BAUTISTA, 1987). x = nombre moyen d'alevins produits par femelle durant une période de 21 jours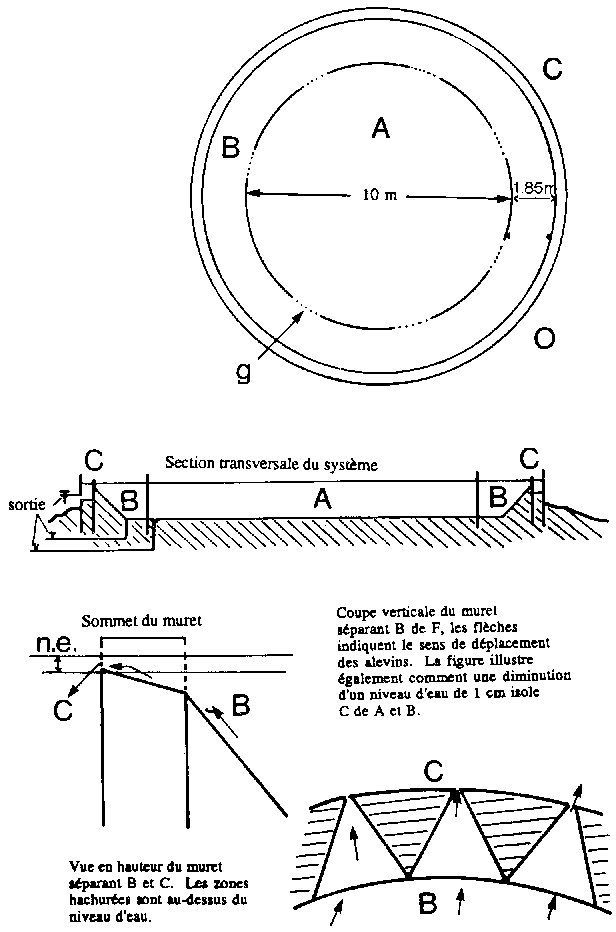{kind=link}
{kind=link}