![]()
![]()
![]()
2.3.3.1. Composition de l'aliment
2.3.3.2. Formulation et taille de l'aliment
2.3.3.3. Ration et fréquence de nourrissage
L'élevage des tilapias en tanks et raceways requiert une alimentation artificielle de qualité, apte à subvenir aux exigences nutritionnelles des poissons en croissance. De nombreux paramètres doivent être pris en considération, tels que la qualité nutritionnelle de l'aliment, en relation avec le type d'alimentation des poissons en milieu naturel (taux de protéines et de lipides, contenu énergétique, proportions de protéines animales et végétales). La granulation et l'appétence, la ration et la fréquence de nourrissage, ainsi que le mode de distribution (manuelle, automatique ou à la demande) sont également très importants.
2.3.3.1.1. Les protéines
Le taux et la nature des protéines
La plupart des travaux concernant la nutrition des tilapias ont tenté de déterminer les besoins en protéines selon les différentes classes de taille. La proportion de protéines dans le régime est en effet de première importance, d'autant plus que les besoins protéiques des poissons sont généralement plus élevés que ceux des animaux terrestres. L'apport en protéines animales dans l'aliment représente souvent plus de la moitié des coûts de l'alimentation (JAUNCEY et ROSS, 1982).
Le niveau de protéines permettant une croissance maximale diminue à mesure que la taille des poissons augmente. Nous présentons, au tableau 2.40, les recommandations de JAUNCEY et ROSS (1982) concernant le taux de protéines nécessaires selon les différentes catégories de tilapias. Pour les larves et les jeunes alevins (<1.0 g), la plupart des auteurs préconisent un régime avec une teneur en protéines avoisinant 50%. HUTABARAT et JAUNCEY (1987) signalent toutefois que le meilleur coefficient de conversion (poids d'aliment distribué gain de poids) est obtenu avec un régime 30% de protéines et un rapport protéines-énergie de 66.69 mg de protéines par kilocalorie d'énergie brute. Des concentrations en protéines inférieures à celles mentionnées, au tableau 2.40 procurent également une croissance satisfaisante. JAUNCEY et ROSS (1982) ont ainsi montré que chez des alevins de T. mossambica de 0.5 à 10 g, le taux optimal de protéines avoisinait 35 à 40%, mais qu'une croissance égale à 80% ou 64% de la croissance maximale était obtenue avec un aliment respectivement à 24 et 16% de protéines. Ceci démontre la possibilité d'utiliser des taux de protéines sub-optimaux pour la croissance, mais éventuellement très intéressants sur le plan économique, en réduisant le coût de l'alimentation.
Chez les géniteurs, il semblerait qu'un régime de 27.5 à 35% de protéines soit optimal pour la croissance. En ce qui concerne la capacité reproductrice (fréquence de ponte, taille des oeufs, fécondité et taux d'éclosion), les résultats de WEE et TUAN (1988) tendent à démontrer que les régimes à teneur faible ou moyenne en protéines (20,27.5,35%) favorisent une fécondité élevée, des pontes tardives et fréquentes et des oeufs de petite taille, alors qu'un régime riche en protéines (42.5 et 50%) permet des pontes plus précoces, mais moins fréquentes.
Digestibilité des protéines et composition en acides aminés
Les teneurs préconisées au tableau 2.40 sont basées sur une digestibilité optimale des protéines, c'est-à-dire supérieure à 90%. Or, les protéines d'origine végétale (P.V.) sont moins assimilables que les protéines animales (P.A.). C'est pourquoi elles devront être inclues à des concentrations plus élevées lorsqu'une fraction importante de celles-ci est d'origine végétale, permettant ainsi une assimilation similaire à celles d'origine animale dont la digestibilité est supérieure à 95%. Les coefficients de digestibilité des protéines, lipides et glucides, ainsi que la valeur énergétique de différents ingrédients pour T. nilotica sont présentés en annexe 2.1.
Chez les tilapias, l'importance du rapport protéines animales/protéines végétales (PA/PV) varie selon les auteurs. Ainsi, chez T. nilotica, HASTINGS (1973) suggère que dans un régime à 30% en protéines, les P.A. ne sont pas essentielles en élevage intensif en étang, alors que SITASIT et SITASIT (1977) obtiennent les résultats inverses. MELARD et PHILIPPART (1981) signalent également qu'avec des aliments à faible teneur en protéines (23-26%), les taux de conversion de l'aliment sont d'autant plus bas que la proportion de P.A. par rapport aux P.V. est plus élevée. Chez d'autres espèces de tilapias tels que T. aurea, T. rendalli et T. mossambica, l'incorporation des protéines animales augmente fortement la digestion et les performances de croissance (GOLDSTEIN, 1970; MILLER, 1976; MATHAVAN et PAUDIAN, 1976; DAVIS et STICKNEY, 1978). En général, le remplacement des P.A. par des P.V. à concurrence de 25% apparaît acceptable (STICKNEY, 1986). APPLER er JAUNCEY (1983) ont toutefois montré qu'un remplacement des protéines animales par des algues (Cladophora glomerata) entraînait une diminution de croissance et d'utilisation des protéines lorsqu'il dépassait 5%.
Tableau 2.40. Proportions des composés alimentaires (en % de la matière sèche) pour différentes catégories de tilapias (d'après JAUNCEY et ROSS, 1982).
|
Composés (% mat. sèche) |
<0.5g |
0.5-10 g |
10-35 g |
35g |
Géniteurs |
|
Protéines brutes |
50 |
35-40 |
30-35 |
25-30 |
30 |
|
Lipides bruts |
10 |
10 |
6-10 |
6 |
8 |
|
Hydrates de C digestibles |
25 |
25 |
25 |
25 |
25 |
|
Fibres |
8 |
8 |
8-10 |
8-10 |
8-10 |
|
Vitamines |
2 |
2 |
2 |
2 |
2 |
|
Minéraux |
4 |
4 |
4 |
4 |
4 |
Les besoins quantitatifs en acides aminés essentiels de T. nilotica sont présentés en annexe 2.1. Ces 10 acides aminés essentiels, communs à toutes les espèces chez qui les besoins ont été déterminés, ne peuvent être synthétisés par le poisson et doivent donc se trouver intégralement dans l'aliment.
Les besoins en acides aminés soufrés (méthionine et cystine) peuvent probablement être couvert soit par la méthionine seule, soit par une combinaison de méthionine et cystine (LIM, 1989).
Sources de protéines
Un inventaire des différentes sources de protéines végétales et animales destinées à l'élevage des tilapias a été réalisé par JAUNCEY et ROSS (1982). Nous présentons ici les données essentielles se référant au nourrissage des alevins, principalement de T. nilotica.
Parmi les protéines d'origine végétale, les plus prometteuses, en tant qu'alternatives aux farines de poisson, sont celles issues de tourteaux d'oléagineux. La plupart sont d'origine tropicale et la concentration en protéines est relativement élevée, de l'ordre de 20 à 50%. Ce sont les tourteaux de soja, coton, arachide, colza, tournesol, sésame, noix de coco et palmier.
PANTHA (1982, in JAUNCEY et ROSS, 1982) a montré que chez les alevins de T. nilotica, aucune différence de performance de croissance et d'utilisation de l'aliment n'était observée entre un régime à 40% de protéines uniquement d'origine animale (farine de hareng) et un régime isoprotéique où 75% de la farine de hareng étaient remplacés par du soja enrichi avec de la méthionine. La méthionine ne doit toutefois pas être additionnée sous sa forme cristalline, mais liée au préalable à la pâte de soja (TESHIMA et KANAZAWA, 1988). De même, OLUKUNDE (1982, in JAUNCEY et ROSS) signale que la croissance des alevins de T. nilotica n'est pas affectée, dans un régime à 45% de protéines, par le remplacement de 11% de la farine de poisson en farine d'arachide et 11% en farine de tournesol. Avec un régime moins riche en protéines (30%), le remplacement des 2/3 de la farine de poisson par du tourteau d'arachide ne permet que 60% des performances de croissance des "fingerlings" de T. nilotica.
L'ensemble des informations disponibles sur l'emploi des tourteaux d'oléagineux indique que ces aliments de substitution ne peuvent remplacer complètement l'apport de protéines animales, mais qu'ils peuvent cependant se révéler efficace en tant que substituants partiels permettant une diminution sensible du coût de l'aliment
En effet, l'utilisation des protéines animales est considérée comme très onéreuse du point de vue économique, principalement dans les pays tropicaux où il existe une carence en protéines animales et où le prix de vente du poisson ne permet pas l'apport d'un aliment coûteux. Idéalement, elles ne devraient contribuer à l'apport en protéines qu'en tant que composant évitant une déficience en certains acides aminés.
Les protéines animales incluent les farines de poisson, de crustacés, de viande ainsi que des sous-produits tels que farine d'os, de sang, de poudre de lait, etc.
2.3.3.1.2. Lipides et glucides
JAUNCEY et ROSS (1982) signalent qu'en terme de kilocalories, les protéines constituent la source d'énergie la plus onéreuse. Le but d'un aliment pour poissons est donc de maximiser l'utilisation des protéines pour la croissance en fournissant une quantité adéquate d'énergie sous forme de lipides et d'hydrates de carbone (action d'épargne des protéines).
Toutefois, à des niveaux modérés de protéines, la quantité d'énergie ne peut dépasser un certain seuil sans provoquer des dépôts de graisse et des changements indésirables dans la composition de la chair du poisson.
Les lipides constituent la première source d'énergie, le contenu énergétique d'un g de lipides (9.1 kcal d'énergie brute) étant deux fois plus élevé que celui d'un g de protéines (5.5 kcal) ou d'un g d'hydrate de C. (4.1 kcal) (JAUNCEY et ROSS, 1982; NEW, 1987). Il semblerait toutefois que les tilapias n'utilisent pas les taux élevés de lipides aussi efficacement que le font les salmonidés ou les cyprins (JAUNCEY, 1979; STICKNEY, 1986), mais peu d'études se sont intéressées à l'utilisation des lipides par ce groupe.
Les lipides servent également de source en certains acides gras essentiels. Afin de satisfaire avec certitude les besoins en acides gras essentiels des tilapias, JAUNCEY et ROSS (1982) recommandent d'inclure dans le régime, 1% de chacune des séries d'acides gras w -3 et w -6. Les besoins semblent toutefois importants pour la famille des acides linoléiques (w -6). Des additions d'huiles végétales (soja ou maïs) riches en 18: 2w -6 procurent des meilleurs taux de croissance que des additions d'huiles de poisson riche en acides gras 20: 5w -3 (TAKENCHI et al, 1983a). Le niveau alimentaire optimal en acides gras w -6 a été estimé à 0.5% chez T. nilotica (TAKENCHI et al, 1983b). Comme l'indique le tableau 2.40, la quantité de lipides à incorporer dans le régime reste relativement constante durant la croissance du poisson jusqu'au stade de juvénile. Un enrichissement en lipides peut éventuellement être effectué dans l'aliment destiné aux géniteurs.
Les quelques études ayant évalué les besoins des tilapias en hydrate de carbone ont montré qu'il existait des possibilités importantes d'épargne des protéines par utilisation d'hydrates de carbone (ANDERSON et al, 1983). Toutefois, les fibres (polysaccharides complexes constitués principalement de cellulose chez les végétaux) ne sont pas utilisables par les tilapias qui, comme les autres poissons, ne disposent pas de cellulase (STICKNEY, 1976). En étang, ces ingrédients non digestibles peuvent être utilisés comme fertilisants (HUET, 1970), mais en tanks, ils entraînent une augmentation de l'encrassement et de la détérioration de la qualité de l'eau (HAUSER, 1975; BALARIN et HALLER, 1982).
En règle générale, on peut admettre que le rapport protéine alimentaire/énergie diminue à mesure que le tilapia grandit. Ainsi, WINFREE et STICKNEY (1981) ont montré que T. aurea (2.5g) présentait la meilleure croissance avec un régime à 56% de protéines et un rapport énergie digestible/protéine (ED.P) de 8.2 kcal/g de protéine. Par contre, chez les poissons de plus grande taille (7.5g), la croissance était maximale avec un régime contenant 9.4 kcal ED/g de protéine. Chez T. nilotica (1.7 - 7g), la croisance optimale est obtenue avec un rapport ED/P de 8.3 kcal/g pour un régime à 36% de protéine (KUBARYK, 1980).
2.3.3.1.3. Vitamines et minéraux
Certains aliments composés contiennent un supplément vitaminé et minéral appelé prémix. Ces prémix, mis au point pour d'autres espèces ont également donné satisfaction chez les tilapias, bien que certains symptômes de déficience (scoliose par ex.) puissent être observés, principalement suite à une carence en vitamine C. La composition des prémix vitaminé et minéral est présentée en annexe 2.2, d'après les informations de JAUNCEY et ROSS (1982).
Signalons toutefois que la vitamine B12 peut être synthétisée dans l'intestin de T. nilotica et qu'il n'est donc pas nécessaire de l'inclure dans le régime (LOVELL et LIMSUWAN, 1982). Même en cas d'une alimentation artificielle, les prémix vitaminé et minéral doivent être additionnés respectivement à raison de 2 et 4% du poids sec de l'aliment (Tableau 2.40). Ces suppléments ne sont, par contre, pas requis dans les élevages de tilapias en étangs, car, dans ce milieu, ils trouvent ce qui leur faut en vitamines et oligo-éléments (NEW, 1987).
La formulation de différents aliments pour alevins de tilapias est présentée en annexe 2.3, d'après les informations récoltées chez plusieurs auteurs (HASTINGS, 1973; HALLER, 1974; CRUZ et LAUDENCIA, 1978; DAVIS et STICKNEY, 1978; BALARIN et HALLER, 1982; JAUNCEY et ROSS, 1982; NEW, 1987). De façon encore plus marquée que chez les autres espèces, il n'y a pas de formulation standard universellement acceptée, puisque l'intérêt d'un ingrédient particulier sera fonction de son abondance et de sa disponibilité selon les différentes régions. Les différents modes de formulation, de préparation (farine, granulés, pâtes,...), et de conservation de l'aliment ont été revus par JAUNCEY et ROSS (1982) et NEW (1987).
La taille de l'aliment doit être en relation avec l'âge du poisson (Tableau 2.41). Il est souhaitable que le granulé soit compact et ne se désagrège pas de suite au contact de l'eau mais les granulés trop durs sont déconseillés.
Tableau 2.41. Tailles des granulés recommandées pour les différentes classes de taille de Tilapia nilotica
|
Age/poids du poisson |
Taille de la particule (f) |
Références |
|
Larve: premières 24 h |
En solution |
MACINTOSH et DE SILVA, 1984 |
|
Larve: 2ème jour -10ème jour |
500 m m |
|
|
Larve: 10ème jour - 30ème jour |
500-1000 m m |
|
|
Alevin: 30ème jour - juvénile de 0.5 à 10g. |
500-1500 m m |
|
|
Alevin: de 1 à 30g. |
1-2 mm |
JAUNCEY et ROSS, 1982 |
|
>30g. |
2-4 mm |
|
La ration journalière peut varier en fonction de plusieurs facteurs tels que la taille du poisson, ses besoins en protéines et sa capacité de digestion, la qualité de l'aliment, les conditions physico-chimiques du milieu (t°, O2, lumière,...),...
En conditions d'élevage intensif (tank et raceway à densité élevée de stockage), on favorisera l'apport d'un aliment aussi riche que possible, - en considérant toutefois le rapport qualité-prix de l'aliment et sa rentabilité financière - ce qui permettra de réduire au maximum la quantité à distribuer et la fraction non digestible (et donc la dégradation du milieu d'élevage). A titre de référence, nous présentons, à la figure 2.11, les rations journalières recommandées, selon la taille des alevins, par SARIG et MAREK (1974) et MELARD (1986). D'autres valeurs ont également été proposées par MELARD et PHILIPPART (1981), BALARIN et HALLER (1982) et NEW (1987). La ration est dite "de maintenance" lorsque la vitesse de croissance est nulle, et "maximale" lorsque la croissance est également maximale. La ration optimale est celle permettant une croissance maximale par unité de ration (figure 2.11).
La tendance naturelle des tilapias à se nourrir de façon relativement continue durant la journée indique que leur système digestif est plutôt adapté à recevoir un apport régulier et fréquent de petites quantités d'aliments. Bien que la fréquence de nourrissage soit également affectée par la taille du poisson et par la température de stockage (agissant sur la vitesse d'évacuation de l'estomac), les auteurs s'accordent à penser que le tilapia doit être nourri peu et souvent.
La fréquence de nourrissage des larves et des jeunes alevins de T. nilotica doit être au minimum de 4 fois par jour et idéalement de 8 fois/jour chez les "fingerlings" (JAUNCEY et ROSS, 1982; NEW, 1987). PIPER (1982) recommande une fréquence de nourrissage telle que 1% de la biomasse soit distribuée à chaque repas et que 90% de la quantité distribuée soit consommée dans les 15 minutes qui suivent sa distribution. Un nourrissage fréquent favorisera également l'obtention d'alevins de taille uniforme. L'utilisation de nourrisseurs automatiques (fréquence de nourrissage élevée) limite considérablement le gaspillage, mais la compétition inter-individuelle pour arriver au point de nourrissage et la dépense d'énergie qui en découle entraîne une diminution de la croissance quand la densité augmente (PETIT 1980, in MELARD et PHILIPPART, 1981).
En conclusion, dans les conditions de production intensive en tanks, raceways ou arènes, la ration alimentaire optimale quotidienne sera élevée pour les petits alevins (11% de la biomasse pour poissons de 2 g), plus faible pour les "fingerlings" (8% pour des poissons de 15 g et 7% pour des poissons de 30 g) et nettement plus faible pour les adultes (3% pour poisson de 150 g). La fréquence de nourrissage manuel compte tenu de l'ensemble des facteurs de production (notamment le coût de la main d'oeuvre) sera optimalisée à 4 distributions bien réparties sur la journée et la dernière distribution sera effectuée au moins deux heures avant la tombée de la nuit. En cas de possibilité, on construira des distributeurs d'aliments automatiques à la demande (cf planche 2.3 et "self-feeding" de HASTINGS, 1973) ou on utilisera les distributeurs programmés d'aliments (type mécanique à ressort ou type solaire). Mais dans ce dernier cas, le niveau de ressources énergétiques et de technicité du pisciculteur devra être nettement plus élevé. Ces données en conditions de croissance optimale (t° = 28°C) sont évidemment à moduler selon la température de l'eau et donc doivent varier selon l'altitude, la latitude et la saison.
Peu d'informations sont disponibles quant à la rentabilité de la production d'alevins de Tilapia nilotica en tanks, car d'une part, ce mode de production intensive est peu développé dans les pays en voie de développement et d'autre part, les bilans économiques considèrent généralement l'ensemble de la production jusqu'au poisson de consommation, et non la production d'alevins seule.
D'après BALARIN et HALLER (1983), l'aliment constitue la part prépondérante (50-60%) des coûts de production en élevage intensif. Les coûts de main d'oeuvre représentent 20%, ceux de pompage 10 à 15% et ceux de transport et d'entretien, 10 à 15% également Ces indications concernent toutefois l'ensemble des coûts de production jusqu'au poisson de consommation.
WRIGHT et KENMUIR (1981) ont établi un inventaire des coûts d'installation et de production d'une pisciculture de Tilapia mossambica au Zimbabwe. A titre indicatif, nous présentons, en annexe 2.5, les points principaux de cette étude se référant aux coûts de reproduction et d'alevinage, jusqu'à un poids moyen de 2 g, pour une pisciculture de 10 ha ayant comme objectif la production de 5 tonnes/ha/an de poissons de consommation.
Le grossissement s'effectue en étang, à une densité de mise en charge de 13250 poissons de 2 g par ha atteignant, à la vidange, un poids moyen de 400 g.
Le coût de l'aliment représente 24% des coûts de production et le coût du personnel 12%. Le coût de production de l'alevin de 2 g, soit 0.012$, équivaut à 4% du prix de vente du poisson de consommation (400g), soit 0.75 $/kg.
Indépendamment des critères purement économiques, d'autres paramètres doivent être pris en considération lors de l'installation d'une pisciculture de T. nilotica en conditions intensives:
les facteurs socio-économiques: l'élevage des tilapias est une activité récente, surtout dans les pays tropicaux. Un transfert de technologie est donc indispensable. Le manque de qualification de la main d'oeuvre représente sans doute un handicap majeur au développement de l'élevage intensif des tilapias en Afrique. Les résultats peuvent s'avérer désastreux si une formation et un encadrement du personnel n'est pas assuré. De même, la disponibilité et le coût des matériaux de construction et de l'aliment déterminent la rentabilité économique d'une installation.le site et les infrastructures existantes: l'élevage intensif nécessite de l'eau en quantité et de qualité élevée, d'autant plus que les densités de stockage sont généralement importantes. Le site doit donc être sélectionné en conséquence. La présence d'une infrastructure de travail (électricité, moyens de communication, machines,...) facilite les opérations de construction et de production (BALARIN et HALLER, 1983).
Comparé à l'alevinage en étangs, l'élevage des tilapias en tanks "raceways" et arènes, par l'aspect généralement intensif qu'il sous-entend, présente un certain nombre d'avantages similaires à ceux relatifs à l'élevage d'autres espèces de poissons tels que salmonidés, cyprinides, à savoir:
1. Production élevée par unité de surface et de volume.2. Possibilité de réaliser la production de différentes phases d'élevage de façon concommittante.
3. Contrôle aisé et permanent des différentes phases de l'élevage et possibilité d'intervention dès le moindre problème.
4. Facilité de traitement thérapeutique lors de l'apparition de maladies.
5. Récolte aisée et fréquente de la production.
6. Calibrage régulier, favorisant la production de poissons de taille uniforme.
Ces arguments, valables quelle que soit la phase de grossissement, sont également applicables à la production de larves et d'alevins. Nous avons en effet montré que T. nilotica se reproduisait spontanément et fréquemment en tank, la production pouvant atteindre jusqu'à 2700 larves/m2/mois (tableau 2.36). Les problèmes inhérents au comportement territorial des mâles et à l'aggressivité des poissons ne participant pas à l'acte sexuel peuvent également être fortement réduits par des installations du type des "arènes" de HALLER et PARKER (1981), bien que les tanks de conception plus simple et de construction moins coûteuse aient donné satisfaction quant à la production de larves.
Le contrôle permanent des géniteurs stockés en tank facilite la récolte régulière des larves à mesure qu'elles sont produites.
La production de T. nilotica depuis le stade de larve de quelques jours (0,02g) jusqu'au stade d'alevin de 30 g, est généralement obtenue avec un taux de mortalité très faible, de l'ordre de 3% (MELARD et PHILIPPART, 1981). Enfin, la croissance de T. nilotica en tank est très rapide, surtout si les alevins sont placés dans des conditions optimales d'alimentation (nourriture riche en protéines) et de qualité d'eau (renouvellement important) ce qui conduit à une production pouvant atteindre 22.5 kg/m2/mois (Tableau 2.39).
Les principaux inconvénients de la production d'alevins de T. nilotica en tanks, raceways ou arènes sont liés à la dépendance totale vis-à-vis de l'aliment artificiel, à l'emploi d'un personnel qualifié et aux coûts des infrastructures et de leur maintenance, pour maintenir une bonne qualité du milieu lors d'élevage à hautes densités.
Comme nous l'avons déjà signalé précédemment, une croissance satisfaisante ne peut être obtenue qu'au prix d'un apport d'aliment composé de qualité souvent importé, rarement produit localement, mais toujours très onéreux. Bien que des ingrédients de substitution riches en protéines végétales (ex: farine de soja) puissent être incorporés, la proportion de protéines d'origine animale, sous forme de farine de poisson, de viande ou de sang, contribue, pour une large part, au coût élevé de l'aliment. C'est pourquoi l'élevage intensif de T. nilotica en tank ne peut s'avérer rentable que dans les pays où le prix de vente du poisson est suffisamment élevé. Dans beaucoup de pays tropicaux, le prix de vente de T. nilotica est relativement faible, ce qui compromet sensiblement la production intensive de ce poisson en tank, raceways et arènes avec un coût d'aliment trop élevé, du moins en ce qui concerne la production de poisson de consommation. BALARIN et HALLER (1979) signalent toutefois qu'au Kenya, la vente d'alevins produits en tank permet un rapport de plus de 400% des coûts de production. Il n'empêche qu'en 1989, Baobab farm semblait connaître de très sérieuses difficultés financières.
Indépendamment du coût de l'aliment, son approvisionnement régulier peut également s'avérer problématique, surtout dans certaines régions intertropicales, et entraîner de la sorte des arrêts ou des ralentissements de la production. A cela, il faut ajouter les difficultés de conservation de cet aliment en régions équatoriales à humidité relative et température toujours élevées.
La production intensive d'alevins de T. nilotica en tanks ou raceways nécessite de plus la présence en permanence d'un personnel qualifié ayant acquis une bonne connaissance des différentes phases de l'élevage et de la gestion de la pisciculture (gestion du stock de géniteurs, contrôle des pontes et récolte des larves, nourrissage régulier et entretien des tanks, etc).
Bien qu'il s'agisse d'un poisson résistant parfaitement aux conditions artificielles d'élevage, le stockage en tank à hautes densités favorise le déclenchement et la propagation de maladies risquant de contaminer tout le stock de poisson en élevage et d'entraîner des mortalités massives, voir totales, si des mesures thérapeutiques ne sont pas mises en oeuvre dès l'apparition des premiers symptômes.
Enfin la production intensive implique des infrastructures coûteuses (tanks en béton ou en polyester, raceways, arènes en béton) qui ne peuvent se justifier que par une production industrielle avec apports importants de capitaux (souvent étrangers). De plus, ces systèmes de production intensive nécessitent un système de pompage et d'aération complémentaire ce qui exige une source d'énergie, l'achat d'un matériel coûteux (pompes électriques, filtres, tuyaux, vannes, etc...) qui est souvent importé et pour lequel se pose le problème d'achat en devises étrangères ainsi que la fourniture de pièces de rechange. Ces élevages industriels (Baobab farm à Mombasa, Avaria au Niger, pisciculture industrielle à Brazzaville, pisciculture en raceways de Ban fora au Burkina Faso) ont connu deux problèmes majeurs: irrégularité de l'approvisionnement en aliments de qualité, et prix de revient trop élevés pour le marché local. En conclusion, c'est sans doute l'ensemble de ces inconvénients qui a, jusqu'à présent, empêché un développement durable de la pisciculture intensive en Afrique. Mais le problème fondamental réside dans les coûts de production trop importants ce qui conduit à des prix de vente du tilapia trop élevés par rapport au faible pouvoir d'achat de la masse des consommateurs africains.
La reproduction et l'alevinage en tanks, raceways et arènes correspond à un niveau d'intensification qui ne se justifie qu'en pisciculture industrielle et est donc hors de portée de l'activité rurale traditionnelle. Les coûts de ce type d'élevage ne peuvent se justifier que si le prix de vente du tilapia est très élevé.
La reproduction sera effectuée de préférence dans le système le plus simple: les tanks circulaires d'un diamètre de 3 à 5 m avec en périphérie des moellons installés tous les 70 cm pour délimiter le territoire des mâles. Une densité de géniteurs de 5 ind/m2 avec un rapport des sexes  /
/ de 3/1 devrait donner de bons résultats.
de 3/1 devrait donner de bons résultats.
La croissance des larves récupérées régulièrement par vidanges partielles des tanks de reproduction sera effectuée en tanks successifs de grossissement avec recalibrage régulier des alevins et diminution progressive de la densité de stockage de 4000 larves/m2 dans des bacs peu profonds (20 à 30 cm d'eau) à 1600 ind de 1g/m3 puis 1000 individus de 10g/m3 jusqu'au stade fingerlings (20 à 30g).
Depuis quelques années, le développement des Facultés des Sciences et des Sciences Agronomiques dans les universités africaines a amené un certain nombre d'étudiants et de chercheurs à s'intéresser à l'élevage des tilapias. Toutefois, ces jeunes universités ne disposent pas toujours d'infrastructures adéquates pour effectuer des recherches fondamentales et appliquées sur ces poissons. Il n'empêche qu'un certain nombre d'hypothèses peuvent être préalablement testées à petite échelle dans des aquariums bien conditionnés. Pour ce faire, il est souvent nécessaire d'assurer sa propre production d'alevins. C'est pourquoi, il nous à semblé intéressant de faire part de notre expérience de production d'alevins de T. nilotica au Centre de Biologie du Comportement et Laboratoire de Pisciculture M. Huet de l'Université Catholique de Louvain (U.C.L.) à Louvain-la-Neuve (Belgique).
Comparé aux autres systèmes d'alevinage de Tilapia nilotica. la production d'alevins en aquarium pose un certain nombre de problèmes. Ceux-ci sont tous, directement ou indirectement, provoqués par le volume réduit qui caractérise ce type particulier de milieu de vie et sont principalement de deux types. D'une part, les conditions de vie eh aquarium (entièrement dépendantes de l'homme) doivent couvrir les exigences physiologiques de l'espèce à reproduire, d'autre part, elles doivent également rencontrer les exigences comportementales qui modulent les interactions entre géniteurs chez cette espèce hautement Sociale. Ces deux aspects, installation générale et gestion des stocks de géniteurs, sont traités séparément et successivement.
2.4.1.1. Types d'aquarium
2.4.1.2. Distribution et épuration des eaux
2.4.1.3. Circuit d'air
2.4.1.4. Eclairage
2.4.1.5. Autres aménagements
2.4.1.6. Entretien des installations
Dans le centre de Louvain-la-Neuve, la salle d'élevage des poissons tropicaux (planche 2.4) est équipée de plusieurs types d'aquariums (d'une contenance de 400 l, de 200 l ou de 70 l) remplissant chacun des fonctions particulières. Ainsi, les aquariums de 4001 (longueur: 200 cm; hauteur: 50 cm; profondeur: 40 cm) servent avant tout au stockage des géniteurs (poissons de 50 à 200 g) mais sont, dans certaines circonstances, également utilisés pour la reproduction. Les aquariums de 200 l (longueur: 100 cm; hauteur: 50 cm; profondeur: 40 cm) peuvent également servir au stockage, mais ils se prêtent particulièrement bien à la reproduction de Tilapia nilotica. car ils garantissent aux femelles incubantes ou prodiguant les soins maternels des conditions de vie satisfaisantes, sur un espace minimal.
Par ailleurs, ces aquariums servent au grossissement d'alevins "fingerling" de plus de 5 g. Quant aux aquariums de 70 l (longueur: 60 cm; hauteur: 40 cm; profondeur: 30 cm), ils servent principalement au grossissement des alevins de 0.05 g à 5 g. Ils permettent également le stockage individuel des géniteurs ou l'isolement de poissons blessés.
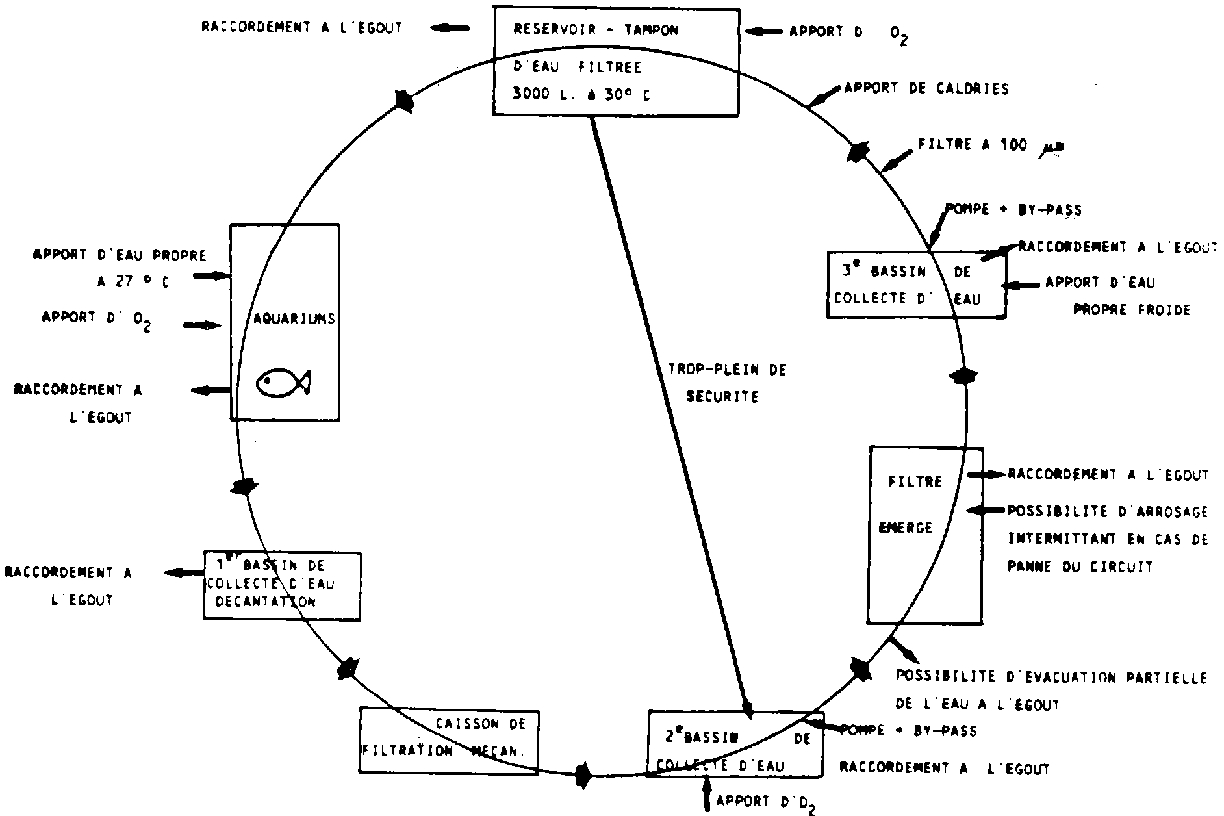
Chacun de ces aquariums peut être alimenté, soit en eau propre, soit en eau recyclée, à bonne température. L'évacuation des eaux usées s'effectue par un trop plein constitué d'un tuyau en PVC (d'un diamètre de 25 mm et d'une hauteur légèrement inférieure à celle de l'aquarium, percé à 4 cm de son bord supérieur d'une rangée de trous, permettant le réglage du niveau de l'eau). L'eau est ensuite envoyée par un tuyau de décharge (diamètre: 32 mm) vers une gouttière (diamètre: 100 mm) qui l'achemine vers le filtre. En cas de maladie, chaque aquarium peut être déconnecté du circuit général et être filtré individuellement.
Dans cette installation (figure 2.12), chaque aquarium est alimenté en dérivation avec de l'eau à bonne température (± 27°C). Ce type de distribution permet de déconnecter un aquarium déterminé en cas de maladie. L'eau des aquariums est renouvellée en permanence à faible débit (0.1 l/min). Selon les cas, elle provient soit de la réserve d'eau propre, soit de la réserve d'eau recyclée, soit encore des deux réseaux de distribution à la fois. Ces réserves sont placées en hauteur et l'eau s'écoule donc par gravité. L'eau excédentaire est évacuée des aquariums par le trop-plein et acheminée par une gouttière vers le filtre.
Celui-ci est constitué de quatre entités bétonnées, chacune étant munie d'un trop-plein et pouvant, si nécessaire, être mise à l'égoût. Le premier compartiment en forme de cône renversé, sert de bassin collecteur et permet une première décantation. En fonction de la quantité d'eau qui afflue vers le filtre, une fine couche d'eau passe ensuite au-dessus du muret qui sépare le premier compartiment du second. L'eau, y traverse d'abord un caisson, rempli de billes d'argex (filtration mécanique) avant d'être pompée vers le filtre biologique du troisième compartiment.
Cette filtration permet la dégradation des produits toxiques contenus dans l'eau (tels l'ammoniaque et les nitrites). Elle s'opère sur des éléments plastiques, appelés garnissage, qui présentent une surface maximale pour le volume occupé. Ces éléments sont empilés les uns sur les autres dans un énorme caisson, placé au-dessus du troisième compartiment. L'eau, en ruisselant sur ces éléments, entre en contact prolongé avec les colonies de bactéries, qui dégradent efficacement les matières organiques. Une filtration biologique maximale est assurée lorsque le filtre biologique a été dimensionné en fonction de la quantité d'eau à filtrer et de sa charge en matière organique et en produits toxiques. Ce rapport entre volume d'eau à filtrer et volume du filtre varie entre 2 (charge en poisson élevée) et 4 (charge en poisson plus faible). Vu le nombre limité de poissons adultes nécessaire à la production d'alevins, le volume du filtre est généralement quatre fois inférieur à celui de l'eau à filtrer. L'eau filtrée est ensuite recueillie dans le bassin du troisième compartiment, d'où elle s'écoule vers le dernier compartiment par un tuyau de connexion.
Dans ce compartiment, une seconde pompe envoie enfin l'eau vers le réservoir (passage obligé par un filtre de 100 microns), situé à l'étage supérieur. Ce dernier joue un rôle tampon. En effet, il assure un apport régulier d'eau vers les aquariums et ce malgré les fluctuations de débit dues à des vidanges d'aquarium, de mise à l'égoût d'un compartiment, etc. Généralement, ce réservoir est rempli à ras-bord et il convient donc d'y installer un trop-plein. Celui-ci peut être raccordé au second compartiment du filtre, évitant ainsi une perte inutile d'eau du circuit.
Dans notre installation, les vitesses d'écoulement vers les aquariums et de repompage vers le réservoir ont été réglées de manière telle que l'eau est brassée 3-4 fois par jour. Néanmoins, vu la faible mise en charge du système, l'eau recyclée n'est remplacée qu'au bout de 4 jours.
En aquarium, l'apport permanent d'oxygène est essentiel à la santé et au bien-être des poissons. Même si Tilapia nilotica résiste relativement bien à une faible saturation en oxygène (< 1 ppm à 25°C), une insuffisance chronique en oxygène réduit sensiblement les fréquences de pontes. Dans notre laboratoire, l'approvisionnement en oxygène est assuré par une turbine (2800 tours/min; 220 volts; 2.3 ampères en triphasé). L'air ainsi produit (débit: 100 m3/h; vitesse: 30 m/sec) est distribué par une conduite en PVC (diamètre: 40 mm; débit: 900 l/h) sur laquelle des prises d'air ont été placées à des intervalles réguliers. Le réglage du débit d'air s'effectue, au niveau des aquariums, par des robinets individuels, l'air étant distribuée par des diffuseurs d'air. L'excédant d'air s'échappe du circuit par un vase d'expansion d'une hauteur d'eau d'1.10 m (niveau supérieur à la hauteur maximale d'eau dans les aquariums), assurant une pression constante dans le circuit. Sauf exception, ce type de circuit fournit aux poissons une eau saturée en oxygène à 70% (5.5-6 ppm d'O2 à 25°C).
Une double rangée de tube néon (lumière blanche, 36 watts), protégée par des caches étanches, et placée à environ 50 cm de la surface de l'eau, fournit la lumière nécessaire (1500 -3000 lux à la surface). L'éclairage étant commandé par une minuterie, un cycle d'éclairement de 12 h de lumière et de 12 h d'obscurité est imposé aux poissons. En effet, Tilapia nilotica est relativement sensible à l'éclairement et recherche activement des zones plus claires. Une intensité de lumière trop faible ou un cycle trop court peuvent donc également affecter le comportement reproducteur de cette espèce. Un cycle d'éclairement allongé (13 à 14 h de lumière) favorisera la gamétogenèse et améliorera les performances de reproduction.
Outre les aménagements qui permettent de rencontrer les exigences biologiques de Tilapia nilotica (température, oxygène, éclairage, qualité de l'eau, espace disponible), d'autres aménagements visent à optimaliser les conditions de travail dans la salle. Ainsi, il est indispensable de prévoir, à même le sol, une évacuation vers l'égout. Celle-ci est facilitée par un sol en légère pente et peut encore l'être davantage par une gouttière enfoncée dans le sol et recouverte d'un grillage qui traverse partiellement ou entièrement la pièce.
L'eau et l'humidité permanente de ces salles impliquent que le revêtement du sol, les plinthes, les murs et les portes y résistent. Par ailleurs, la sécurité de la pièce peut être sensiblement augmentée en suspendant les installations électriques (éclairage, prises électriques) à des rampes disposées au-dessus des aquariums, le danger d'électrocution étant écarté par des prises de terre et des disjoncteurs différentiels. Souvent, un système de ventilation s'avère également nécessaire pour réduire le taux d'humidité de la pièce et pour empêcher la formation de moisissures. En régions tempérées, un double vitrage s'indique pour réduire les pertes de calories et pour éviter la condensation sur les vitres dans le cas où la salle est équipée de fenêtres. Pour l'élevage des poissons, l'absence de lumière naturelle ne pose néanmoins aucun problème. Au contraire, elle permet d'éviter la prolifération des algues vertes.
L'entretien des installations est fort aisé et demande relativement peu de temps. Ainsi, les aquariums nécessitent un entretien toutes les 1-2 semaines. Celui-ci consiste en un nettoyage des parois et une vidange aux trots-quarts, permettant l'évacuation des déchets, sans que les poissons ne soient manipulés, le nouveau remplissage se faisant automatiquement.
Quant au filtre, l'entretien régulier se limite à la vidange hebdomadaire du tank de décantation (5 min) et au nettoyage mensuel du caisson à billes d'argex, des préfiltres et crépines (3 heures). Par précaution, nous contrôlons également toutes les semaines la qualité de l'eau du circuit de recirculation (température, pH, taux d'oxygène, taux de nitrites avant et après filtration, durée 1 h).
Tous les 2-3 mois, le filtre (100 microns) est rincé et les tuyaux de distribution d'eau (diamètre: 10 mm) au-dessus des aquariums débouchés. Enfin, les tanks en béton qui récoltent l'eau du filtre sont brossés et purgés au bout de 6 mois, le nettoyage des réservoirs d'eau ne s'avérant nécessaire qu'après 3-4 ans. Selon d'autres chercheurs qui utilisent le même type de filtre émergé, l'entretien de ce dernier ne s'impose qu'au bout de 10-12 ans et s'annonce par une augmentation notable de sa consommation en oxygène. Sa durée d'activité est donc particulièrement longue.
Néanmoins, un certain nombre de précautions doivent être prises pour lui assurer des conditions de fonctionnement optimales. Etant donné que la filtration se fait de manière biologique, toute action qui risque de détruire la colonie bactérienne doit absolument être évitée. Ainsi, il faut à tout prix s'abstenir d'utiliser dans le circuit (aquariums, gouttière, tuyauterie) des produits toxiques (eau de Javel, permanganate de potassium, antibiotiques etc.), étant donné qu'ils seront directement acheminés vers le filtre. En outre, il faut également s'assurer que le filtre émergé garde, à tout moment, un taux d'humidité élevé, qui est indispensable au bon fonctionnement et à la survie des bactéries. Il arrive parfois que des problèmes surviennent dans l'acheminement de l'eau vers le filtre (pompe en panne, coupure du circuit d'eau pour raison d'entretien etc.). Dans ce cas, il est impératif d'arroser le filtre avec de l'eau propre (qui peut même être plus froide) le temps nécessaire à la remise en état du circuit. De cette manière, il est possible de sauvegarder le pouvoir filtrant du filtre, même si le circuit a été arrêté pendant plusieurs semaines. A l'occasion d'une telle interruption prolongée, nous avons, en effet, constaté que, grâce à cet arrosage compensatoire, le pouvoir filtrant avait entièrement été rétabli après 4-5 jours, alors qu'il aurait fallu plus de 4 semaines pour relancer le filtre après un dessèchement total. Dans le cas où le filtre ne travaille que partiellement, il est conseillé de le charger le moins possible en matière organique, ce qui revient à nourrir moins abondamment les poissons.
2.4.2.1. Maintenance des géniteurs
2.4.2.2. Gestion des géniteurs pour la production d'alevins
2.4.2.3. Elevage de la descendance
Le contrôle complet du cycle de vie de Tilapia nilotica est relativement aisé en laboratoire, à condition d'éviter un certain nombre de difficultés qui sont essentiellement d'ordre pathologique (Annexe 2.6) et comportemental. Celles-ci sont liées, chez les géniteurs, au comportement de défense territoriale (qui se traduit par une agressivité excessive) et, chez les alevins, au comportement cannibale. Ces problèmes trouvent leur origine dans le bouleversement profond que subit l'organisation sociale de cette espèce adaptée aux grandes surfaces aquatiques, lors du confinement spatial en aquarium.
En laboratoire, il est utile de disposer, pendant toute l'année, d'un stock réduit de géniteurs qui permet de relancer la production de poissons en cas de besoin. Idéalement, un stock minimal doit être constitué de 5-6 mâles et de 10-12 femelles (poids des géniteurs entre 100 et 200 g) et peut être gardé dans un seul aquarium de 400 l. Un tel stock, dont les potentialités reproductives dépassent largement les besoins moyens de tests en laboratoire (une femelle de 150 g produit sans problème 300 alevins par ponte), se justifie par le souci de garantir une production d'un millier d'alevins en un mois et ce malgré une baisse momentanée de la fécondité du stock, due à des blessures chez une partie des mâles et/ou à un allongement imprévu du cycle de ponte chez une partie des femelles.
L'évitement des conséquences néfastes de l'agressivité est aisé en dehors de la reproduction. Pour ce faire, il suffît de regrouper les adultes à une densité relativement élevée. En effet, dans ces conditions, les perturbations mutuelles entre congénères rendent difficile la défense territoriale, qui motive pour une part importante l'agressivité des mâles. Il a été démontré qu'en période de conditions défavorables, l'espèce abandonne son investissement dans la reproduction et adopte une vie grégaire, pacifique (FALTER, 1984; FALTER et DE BACKER, 1988). En dessous de 20°C, l'activité sexuelle est totalement arrêtée, mais l'appétit des poissons (et donc leur croissance) diminue également. Enfin, un dernier moyen pour contrôler l'agressivité des mâles consiste à installer un courant d'eau sur le fond de l'aquarium. En tous cas, la pratique courante qui consiste à retirer de l'aquarium l'animal le plus agressif est à proscrire, car elle favorise chez les poissons restants l'apparition de comportements territoriaux. Ces conditions légèrement défavorables à l'espèce (même pratiquée sur plusieurs semaines ou mois) n'handicapent en aucune manière son pouvoir reproductif, à condition que les animaux reçoivent une nourriture en quantité et qualité suffisante et qu'ils ne soient pas trop vieux (âge optimum: 6-18 mois). Dans ces conditions de stockage, la reproduction n'est pas inhibée, mais sa fréquence simplement ralentie. Si les reproductions spontanées doivent être totalement supprimées, il est indispensable de séparer les sexes. Mais même dans ces conditions, certaines femelles continuent à pondre. Elles n'incuberont cependant que pendant 2-3 jours, le temps nécessaire aux oeufs non fécondés de se dégrader dans leur cavité buccale (ROTHBARD, 1979).
Avant de procéder à la description concrète des techniques de reproduction, il faut souligner que le taux de consanguinité croît rapidement avec chaque nouvelle génération vu la fréquence élevée des reproductions d'un petit nombre de géniteurs. En laboratoire, les croisements répétés entre individus apparentés sont toutefois pratique courante et ne semblent pas affecter la viabilité des poissons, à condition que la souche de départ soit de bonne qualité (pas de déformations dans la descendance, bonne croissance et résistance aux maladies, rythme soutenu de reproduction). Dans notre propre souche, nous avons pu constater qu'après quinze générations de reproduction entre individus apparentés, la part de la variance génétique dans la variabilité générale est devenue nulle pour les caractères "agressivité" des mâles (FALTER et DE JAEGERE, 1989) et "rythme de ponte" des femelles (DUFAYT, 1988).
Selon le but recherché et la disponibilité des chercheurs, trois techniques de reproduction peuvent être envisagées: reproduction et incubation naturelles de couples ou de groupes de géniteurs et reproduction artificielle.
2.4.2.2.1. Reproduction et incubation naturelles par couples
Cette technique, qui permet d'obtenir par voie naturelle une progéniture dont le père et la mère sont connus, repose sur le choix sélectif des poissons et leur réunion juste avant la reproduction. Elle nécessite une surveillance soutenue de l'état physiologique du stock des géniteurs, le repérage des individus prêts à se reproduire se faisant sur base de critères parfaitement visibles. Ainsi, les mâles qui défendent un territoire de reproduction, arborent des livrées de colorations typiques (couleur claire et brillante du corps, absence de stries bordant la nageoire dorsale, papille génitale proéminente) (FALTER, 1987) et présentent des comportements caractéristiques (défense d'une aire sur le fond, attaque des congénères, creusement) (HEINRICH, 1968). Les femelles prêtes à pondre deviennent, elles aussi, claires et brillantes, plus agressives et ont une papille génitale protubérante. Lors de l'approche de l'oviposition, elles recherchent activement la présence d'un mâle territorial (alors qu'elles se tiennent d'habitude à l'écart) et se mettent également à creuser (ROTHBARD, 1979).
Chez les tilapias, la vision d'un congénère favorise la maturation gonadique dans les deux sexes. C'est la raison pour laquelle les poissons sont généralement gardés en groupe (sauf en cas de blessure où il vaut mieux les isoler). La première étape dans la préparation des géniteurs consiste à placer deux mâles matures, séparés l'un de l'autre par une cloison solide, transparente et amovible (ex.: moustiquaire sur cadre rigide, retenue par des glissières) dans un aquarium de 200 l. Les poissons y établissent leur territoire en quelques heures. Généralement, ces préparatifs se font le matin, étant donné que Tilapia nilotica se reproduit principalement l'après-midi et le soir (FALTER, 1986). Dès qu'une femelle est prête à pondre, l'un des deux mâles est enlevé et la femelle introduite dans le compartiment libre. Celle-ci peut ensuite s'habituer à son nouvel environnement pendant quelques temps (déterminé par la durée nécessaire à la réapparition de comportements et patrons de coloration sexuels). Pendant cette période d'attente, le mâle et la femelle communiquent visuellement et olfactivement, et se stimulent mutuellement (ARONSON, 1945; SILVERMAN, 1978 a, b, c; FALTER et DOLISY, 1989; FALTER et CHARLIER, 1989). Dès la réapparition des comportements sexuels chez la femelle, la cloison est enlevée et les géniteurs se réunissent spontanément.
En général, le mâle commence directement sa parade devant la femelle et l'invite à le suivre vers son territoire. La femelle ne l'accompagne cependant pas nécessairement dès le début. Dans cette situation, les géniteurs échangent normalement peu de comportements agressifs. Toutefois, si l'un des deux poissons se met à agresser son partenaire (au point que ce dernier perde ses colorations typiques et commence systématiquement à fuir), il est préférable de séparer à nouveau les géniteurs et de présenter un autre mâle (plus pacifique ou plus dominant selon le cas) à la femelle. A cet égard, il est important de donner à la femelle, un mâle d'un poids légèrement supérieur au sien.
Une fois la ponte et la fécondation achevée, il est souhaitable de séparer à nouveau les poissons. C'est à ce stade que le mâle est généralement retiré de l'aquarium et qu'une autre femelle y est introduite pour tenir compagnie à la femelle incubante. Pendant l'incubation (qui dure environ 11 jours à 27°C), la femelle réagit, en effet, plus violemment aux perturbations extérieures et risque de détruire sa progéniture. Il est donc impossible de l'isoler, de la manipuler ou de la pêcher dans cet état. Le transport est néanmoins possible, à condition de ne pas la sortir de l'eau (ex.: transport dans un petit aquarium plongé dans son aquarium de séjour). En outre, il faut savoir qu'une femelle qui a recraché ses oeufs, ne les reprendra plus en bouche.
En général, la femelle garde ses jeunes encore pendant plusieurs jours (2-7 jours) après la phase d'incubation. Au début de cette période, elle les rappelle régulièrement en adoptant une posture particulière (corps incliné à 30%, tête vers le bas) et en battant énergiquement de ses nageoires pectorales. A ce signal, les jeunes refluent vers la tête de leur mère et cherchent, par toutes les voies (y compris les fentes branchiales) un accès à sa bouche. Par la suite, elle ne les rappelle plus qu'en cas de danger. Dans la nature, les jeunes se séparent de leur mère après quelques jours. En aquarium, la mère se désintéresse de ses jeunes au bout d'une semaine. C'est à ce moment qu'il est le plus indiqué de séparer la mère de ses jeunes. Elle pourrait tout au plus encore rester une semaine avec ses jeunes. En effet, la séparation doit absolument survenir avant que la femelle ne se prépare à une nouvelle ponte, étant donné qu'après cette oviposition, elle reprend en bouche tout ce qu'elle trouve sur son chemin. C'est ainsi que nous avons un jour inspecté la bouche d'une femelle qui contenait, outre les oeufs fraîchement pondus, une partie du gravier et une cinquantaine d'alevins de la ponte précédente. Depuis cet incident, tous nos aquariums sont dépourvus de gravier (ou de sable). Cette absence de substrat permet également d'assurer une meilleure hygiène de l'aquarium et n'est nullement indispensable au bon déroulement de la reproduction.
2.4.2.2.2. Reproduction et incubation naturelles par groupes
Cette seconde technique s'inspire du système d'élevage développé par HALLER et PARKER (1981). Elle consiste à introduire simultanément un mâle et trois à quatre femelles (poissons matures mais pas nécessairement prêts à se reproduire) dans un aquarium de 200 l (ou un mâle et six à dix femelles dans un aquarium de 400 l). Une cloison, solidement fixée à l'aquarium et percée de quelques ouvertures (suffisamment grandes pour permettre le passage aux femelles) retient le mâle territorial et agressif dans son compartiment, alors qu'elle permet aux femelles, plus petites, de circuler à leur gré dans tout l'aquarium. Ce système offre deux grands avantages. D'une part il ne nécessite pas une trop grande surveillance, et d'autre part, il minimise le risque qu'une femelle prête à pondre ne soit fécondée. Le seul inconvénient réside dans le fait que les poissons, une fois prêts, se reproduisent rapidement et que l'observateur humain n'y assiste que rarement.
Dans ce système, le mâle doit être retiré au plus tard la veille du jour où le première femelle fécondée libère ses alevins. La femelle incubante annonce ce moment en arborant un patron de coloration typique, caractérisé par l'apparition de 7-8 stries noires verticales sur le corps et de stries noires horizontales entre les yeux, bordés d'un anneau noir. Il est également impératif de retirer toutes les femelles non incubantes, étant donné qu'elles dévorent les alevins fraîchement libérés. Pendant la garde des alevins (qui peut durer 2 à 10 jours), la femelle défend un territoire et est excessivement agressive (plus agressive qu'un mâle territorial). Il faut donc éviter de la rapprocher trop de tout autre poisson. En effet, une telle présence est ressentie comme une menace et la femelle s'épuise à attaquer l'intrus. Tant que ce danger n'est pas écarté, elle ne lâchera pas ses alevins, qui risquent de mourir dans sa bouche. Lorsque l'on dispose d'un nombre suffisant d'aquarium, il est donc préférable de réserver un aquarium par femelle incubante. Enfin, ce système donne même de bons résultats dans des aquariums de 100 l.
Une variante de cette technique a été proposée par ROTHBARD (1979). Pour éviter les conséquences néfastes des comportements agressifs, ce chercheur suggère de constituer, avant la maturité sexuelle, des "familles" de Tilapia nilotica. A cet effet, il place une cinquantaine de poissons de 20 g dans un aquarium de 400 l (200 × 50 × 40 cm). Dès qu'un mâle devient mature et territorial, ROTHBARD enlève une partie importante de la population et ne laisse plus qu'une dizaine de femelles auprès de ce mâle. Selon notre propre expérience, il n'est cependant pas nécessaire de prendre autant de précautions. Même s'il est vrai qu'il est dangereux d'introduire un mâle dans un groupe dont la structure sociale est établie (ce qui se fait endéans quelques heures), il est possible de former à tout moment un groupe de géniteurs, à condition que tous les individus soient introduits simultanément dans un nouvel environnement. Par contre, l'intégration d'une femelle dans un groupe existant ne pose jamais de sérieuses difficultés, à condition qu'elle soit d'une taille comparable aux autres femelles.
2.4.2.2.3. Reproduction artificielle
Comparée à l'importance des recherches menées pour maîtriser les différents aspects de l'élevage de Tilapia nilotica, l'effort fourni pour développer une technique satisfaisante de reproduction et/ou d'incubation artificielle a été négligeable. Le manque d'intérêt pour cette dernière dimension de la biologie de ces espèces est essentiellement dû à la facilité avec laquelle ces dernières se reproduisent spontanément. Néanmoins, la maîtrise de la reproduction artificielle s'avère utile pour des études de sélection génétique et d'hybridation.
Préparation des géniteurs
L'achèvement, en temps voulu, de la maturation gonadique constitue la difficulté principale lors de la préparation des géniteurs. Chez les poissons matures, celle-ci est permanente, bien qu'elle soit cyclique chez les femelles. Chez les mâles, la spermatogenèse est continue, à condition que les poissons soient suffisamment nourris (ration journalière de 2% avec un aliment contenant 25 à 30% de protéines animales) et maintenus dans un environnement physique et social, propice à la reproduction.
Néanmoins, un contrôle de motilité et de concentration des spermatozoïdes s'impose dans le cas où l'on ne travaille qu'avec un ou deux mâles. A cet effet, quelques gouttelettes de laitance (liquide blanchâtre) sont prélevées à sec par pression sur l'abdomen d'un mâle territorial. Placées sous le microscope (agrandissement 400 x), leur motilité est ensuite contrôlée en ajoutant quelques gouttes d'eau. En fait, un tel contrôle s'impose chaque fois que la réussite des reproductions dépend d'un nombre très limité de mâles.
Chez les femelles, l'intervalle entre deux pontes est théoriquement de 28 jours. Cette durée peut néanmoins être diminuée de moitié dans le cas où les femelles sont empêchées d'incuber. Etant donné la difficulté avec laquelle l'état d'avancement de l'ovogenèse entre deux pontes est estimé, DADZIE (1970) propose, pour Tilapia aurea, soit une triple injection, par voie intrapéritonéale, de l'hormone gonadotrope chorionique humaine, dont la dose totale équivaut à 25 U.I./100 g de poids vif, soit une triple injection combinant l'extrait hypophysaire de carpe et l'HCG (dose totale: 0.25 mg/100 g et 25 U.I./100 g). Malgré les résultats encourageants de DADZIE (6 femelles/6 ont pondu dans les 24 heures), la plupart des auteurs, désireux de reproduire Tilapia nilotica de manière artificielle, préfèrent recourir à la méthode VALENTI (1975) qui consiste à utiliser une femelle qui vient de déposer spontanément son premier lot d'oeufs dans l'aquarium.
Reproduction artificielle
Pour réussir une reproduction artificielle, il est indispensable de respecter un certain nombre de règles et il est avant tout conseillé de travailler en équipe. Etant donné que toute la séquence des manipulations doit se dérouler dans un minimum de temps, l'expérience doit être entièrement programmée à l'avance. Comme matériel, on utilise généralement un rouleau de papier fin, un récipient sec à fond arrondi, une plume souple, une pipette Pasteur, un chronomètre et un peu d'eau non chlorée (ou de liquide fertilisant à température dé la pièce). Dès la sortie de l'eau, le géniteur est séché (l'eau diminue considérablement la durée de vie des produits sexuels) et partiellement enveloppé dans du papier fin, préalablement mouillé, seule, la région arrière étant dégagée. Cette dernière mesure permet non seulement de protéger les écailles du poisson pendant la manipulation mais assure également une meilleure manutention de l'animal.
Les ovules étant plus résistants que la laitance, on procède d'abord au "stripping" de la femelle. A cet effet, un premier expérimentateur maintient la papille génitale de la femelle à quelques centimètres seulement du fond du récipient. Avec le pouce et l'index, il exerce ensuite une légère pression sur l'abdomen de la femelle en direction de l'oviducte. Un second expérimentateur surveille l'opération et arrête avec un morceau de papier sec toutes les éjections parasites (urines, fèces, écailles). Si la femelle est réellement prête à pondre, l'extraction des oeufs se fait sans aucune résistance et le premier lot d'oeufs (plusieurs dizaines d'oeufs) est expulsé après trois ou quatre tentatives. A chaque nouvelle pression, un autre lot d'oeufs est expulsé. Lorsque la réserve d'oeufs commence à s'épuiser, leur apparition se fait plus rare et il est alors indiqué d'exercer la pression plus en avant, les ovaires de Tilapia nilotica occupant une large partie du bas de la cavité abdominale. Lorsque les oeufs n'apparaissent pas spontanément après quelques pressions, il vaut mieux interrompre la manipulation. En aucun cas, il ne faut augmenter la pression sur le ventre de la femelle ou insister jusqu'à l'apparition de sang.
Une fois les ovules récoltés, on passe immédiatement à la collecte de la laitance. Le premier expérimentateur, après avoir retourné le mâle sur le dos, exerce à nouveau une légère pression sur le haut de l'abdomen du poisson, les testicules étant moins volumineux que les ovaires et localisés plus près de la colonne vertébrale. Comme précédemment, le second expérimentateur recueille séparément les éjections parasites, et aspire par ailleurs, au moyen de la pipette, la laitance qui coule le long du flanc du mâle. Les mâles de Tilapia nilotica étant généralement fort généreux, il arrive fréquemment qu'un mâle donne plus de 0.5 ml de laitance. Afin de disposer d'une quantité suffisante de laitance, plusieurs mâles (2-3) sont cependant strippés par femelle utilisée.
La fécondation se fait alors en plusieurs étapes. CHOURROUT et ITSKOVITCH (1983) conseillent d'apporter initialement 0.05-0.2 ce de laitance par 100 ovules. Ce mélange est ensuite remué en douceur au moyen d'une plume durant 2 minutes environ. Après cette première phase d'homogénéisation, 10 ce de solution physiologique sont ajoutés et l'ensemble mélangé à nouveau pendant 2 minutes. Alors que ROTHBARD et PRUGININ (1975) rincent les oeufs dès ce stade, CHOURROUT et ITSKOVITCH transvasent les oeufs avec leur liquide dans un récipient plus grand, contenant 100 ce d'eau. Ce n'est qu'après une demie-heure, que les oeufs sont rincés et introduits dans un incubateur (planche 2.4). Nos propres essais sur la durée de vie de la laitance de Tilapia nilotica ont montré, qu'en présence exclusive d'une solution physiologique ou d'une simple solution de NaCl (0.9%), la motilité des spermatozoïdes est nulle, que l'apport supplémentaire d'une petite quantité d'eau douée permet de la déclencher et qu'enfin, l'utilisation de différentes solutions fertilisantes augmente sensiblement leur durée d'activité.
Incubation artificielle
L'incubation artificielle d'oeufs de diverses espèces de Tilapia a été pendant longtemps problématique. Les difficultés provenaient essentiellement des particularités qui caractérisent les oeufs d'incubateurs buccaux. Déjà SHAW et ARONSON (1954), dans leur étude sur le développement embryonnaire de Tilapia macrocephala, avaient découvert que les oeufs devaient être brassés en permanence vu l'abondance des réserves vitellines qui risquent d'étouffer le pôle animal. Ces auteurs avaient également remarqué que les oeufs de Tilapia étaient particulièrement sujets à l'envahissement par des champignons et des bactéries. RANA (1986) vient de confirmer ces données. Il a, en effet, réussi à augmenter le taux de survie d'embryons incubés artificiellement en irradiant l'eau de circulation aux rayons ultra-violets.
Par ailleurs, il a également constaté la fragilité relative du chorion qui se déchire prématurément à la suite de chocs mécaniques trop importants. Actuellement, les incubateurs qui donnent des résultats satisfaisants (60-70% d'alevins à vésicule résorbé), tiennent compte de ces différentes caractéristiques.
A ce jour, différentes techniques ont été développées (SHAW et ARONSON, 1954; BAUER, 1968; NUSSBAUM et CHERVINSKY, 1968; MIRES, 1973; ROTHBARD et PRUGININ, 1975; VALENTI, 1975; LEE, 1979; ROTHBARD et HULATA, 1980; RANA, 1986. DON et al, 1987). Leurs résultats sont néanmoins fort variables, comme le montre le tableau 2.42.
Tableau 2.42. Fonctionnement et rendement de différents types d'incubateurs.
|
Espèce |
Reprod. |
T° |
Forme incub. |
Mouvem. d'eau |
Traitem. Eau |
% éclosion |
Remarques |
Auteurs |
|
T. nilotica |
naturelle |
27° |
bouteille |
diffuseur d'air |
- |
- |
- |
Nussbaum et Chervinsky, 1968 |
|
T. aurea |
artificielle |
32° |
cône renversé |
diffuseur d'air dans goulot |
50U.I./ml pénicilline G et 0.05 mg/mide streptomycine (sulfate) |
56% |
traitem. supplém. (choc au froid) |
Valenti, 1975 |
|
T. nilotica |
artificielle |
26° |
bouteille de Zoug (5l) |
arrivée d'eau par le goulot |
- |
80% (74%-97%) |
- |
Chourrout et Itskovitch, 1983 |
|
T. nilotica |
artificielle |
|
bouteille de Zoug |
- |
avec U.V. |
60% (±10.6%) |
filtration sur charbon actif |
Don et al, 1987 |
|
T. nilotica |
naturelle |
26° |
bouteille de 35ml |
- |
- |
71% (53%-83%) |
- |
Hoerstgen-Schwark et al, 1987 |
Malgré les importantes différences qui existent entre les techniques et qui rendent difficile la comparaison des résultats, les travaux récents semblent néanmoins privilégier:
- l'utilisation d'oeufs fécondés, obtenus après reproduction naturelle,- la mise en route de l'incubation artificielle avec des oeufs âgés de deux jours (stade de pigmentation),
- une stérilisation de l'eau par irradiation aux U.V.,
- un brassage lent des oeufs (bouteille à fond arrondi).
Lors de nos propres essais, nous avons remarqué que le séjour des embryons dans l'incubateur pouvait être limité à la période précédent l'éclosion. Celle-ci survient, à 27°C, entre 96-120 heures après la fécondation. Dès l'éclosion, les alevins ont été transférés dans des récipients peu profonds (2 cm de hauteur d'eau), qui ont été alimentés en permanence par un faible débit d'eau. Dès ce stade, les cas de mortalité ont été extrêmement rares (< 1%).
Etant donné que les conditions de vie en aquarium influencent non seulement le comportement des géniteurs mais également tout le développement de leur descendance, il est utile de commencer par préciser certains termes. En aquarium, la maturité sexuelle apparaît nettement plus tôt que dans les autres systèmes d'élevage. Ainsi, il n'est pas rare devoir qu'un mâle, à peine âgé de trois mois et pesant moins de 10 g, défende déjà un territoire de reproduction. Vers 5-6 mois, la majorité des femelles commencent à pondre, alors qu'elles ne pèsent que 25 à 40 g. Il va donc de soi que les termes "alevins", "fingerlings" et "adultes" ont une signification différente selon le milieu de croissance. En aquarium, nous entendons par "alevins" des jeunes poissons dont le poids se situe entre 0.01 g (1er jour de la nage libre) et 5g. Le terme "fingerling" désigne ensuite des poissons juvéniles entre 5-20 g. Vers 20 g, les poissons commencent à devenir sexuellement matures et accèdent donc au stade "adulte".
2.4.2.3.1. Alevinage
Selon que les alevins ont été obtenus par incubation naturelle ou incubation artificielle, la première semaine de croissance se déroule soit en présence de leur mère (dans un aquarium de 200 l) soit dans le récipient peu profond (2 l pour une ponte). En général, les alevins sont exclusivement nourris avec des nauplii d'artémias (éclos dans un récipient profond contenant 21 d'eau, bien oxygénée à 27°C, à laquelle ont été ajoutés une cuillère d'oeufs d'artémias et 66 g de sel de cuisine). Cependant, d'autres nourritures, riches en protéines (50% de protéines) peuvent également convenir.
Après cette première semaine de croissance (tableau 2.43), les alevins (d'un poids moyen de 0.16 g) sont transférés, par groupe de 50, vers des aquariums de 701. Ils y sont nourris ad libitum 5-6 fois par jour pendant un mois. Ayant atteint à ce moment un poids moyen de 2 g, les groupes sont répartis chacun en deux aquariums, ce qui permet aux poissons de poursuivre leur croissance rapide pendant un autre mois.
Tableau 2.43. Schéma d'élevage d'alevins de Tilapia nilotica en aquarium.
|
Jours |
Aquarium (l) |
Densité (ind.) |
Nourriture |
Poids final (g) |
|
0-7 |
200 |
300-400 |
Artémia |
0.16g |
|
8-35 |
70 |
50 |
Granulés réduits en poudre (50% prot.) |
2.0g |
|
36-64 |
70 |
25 |
Granulés réduits (30% prot.) |
6.0g |
C'est au cours de la période d'élevage des alevins que le cannibalisme est le plus à craindre (PANTASTICO et al, 1988). BERRIOS-HERNANDEZ et SNOW (1983) ont chiffré l'étendue de ces pertes dans leur étude comparative sur les taux de survie d'alevins, élevés dans différents systèmes d'élevage. La perte de loin la plus importante (97%) survient dans le système d'élevage dans lequel les alevins sont maintenus en présence des géniteurs. Les pertes les plus faibles ont été signalées pour les systèmes dans lesquels les géniteurs ont été séparés précocement, soit de leurs oeufs, soit de leurs alevins.
Il serait néanmoins erroné de croire que seuls les géniteurs dévorent les alevins. A plusieurs reprises, nous avons en effet observé en aquarium qu'une partie importante d'une ponte disparaissait, alors que la mère défendait âprement et efficacement ses jeunes contre tous les adultes. Dans ces cas, la perte importante était due au cannibalisme des alevins plus âgés ou des "fingerlings" qui étaient bien tolérés par la mère au milieu du nuage de ses jeunes. Il arrive également que le cannibalisme apparaisse parmi les alevins d'une même ponte. Dans ce cas, ce comportement n'apparaît que lorsque la taille des frères et soeurs diverge sensiblement (rapport 3 à 1). Dans le système d'élevage décrit ci-dessus, les pertes d'alevins sont quasi inexistantes, étant donné que tous les individus grandissent de manière plus ou moins comparable.
D'autres facteurs peuvent également être responsables de la perte plus ou moins importante d'alevins. Ainsi, la qualité de l'eau joue un rôle essentiel. Les alevins sont nettement plus sensibles aux nitrites que les adultes. Une eau de mauvaise qualité, non seulement freine gravement et durablement la croissance, mais encore peut causer une mortalité massive par intoxication. En outre, elle favorise l'explosion bactérienne (ex.: Aeromonas spp). Le seul remède à ce fléau consiste à renouveller l'eau très fréquemment. Par ailleurs, HENDERSON-ARZAPALO et STICKNEY (1980) ont mis en évidence l'existence d'une diminution importante des défenses immunitaires chez les jeunes T. mossambica (âgés de 3 mois environ) qui avaient été élevés en aquarium à forte densité (60 poissons/60 l). Dans ces conditions, la mortalité journalière était de 1 à 2%.
Par contre, des pertes de nature plus accidentelle, dues par exemple à une aspiration par la pompe (ou par le trop-plein) ou à des fluctuations excessives de température, pourraient aisément être évitées. Enfin, contrairement aux attentes, le facteur "aliment" ne semble jouer qu'un rôle secondaire dans la mortalité des alevins. Même sous un régime alimentaire totalement inadéquat, subi pendant deux mois, le taux de survie chez les alevins de T. nilotica (dont le poids n'avait varié que de 0.013 g à 0.1-0.2 g) avoisinait toujours les 50% (PANTASTICO et al, 1981). Néanmoins, une telle malnutrition ou sous-alimentation induit inévitablement l'apparition d'une plus grande hétérogénéité dans la taille des alevins et favorise, de ce fait, le développement du cannibalisme.
2.4.2.3.2. Elevage des fingerlings
Dès que les alevins ont atteint 5-6 g, il est nécessaire de leur offrir un volume d'eau plus important, c'est-à-dire de les transférer vers des aquariums de grossissement. Généralement, nous plaçons 25 poissons par aquarium de 2001. Ils sont ensuite nourris quotidiennement avec une ration alimentaire qui correspond à 3% de leur biomasse. Deux mois plus tard, les premiers signes de maturité sexuelle apparaissent chez les mâles, alors qu'ils ne le feront qu'un mois plus tard chez les femelles.
![]()
![]()
![]()
{kind=link}