![]()
![]()
![]()
H.I. Joly1
Laboratoire INRA-ENGREF de sciences forestières
Unité de génétique des populations d'arbres forestiers
associée au CIRAD-Forêt, France
Adresse actuelle: CIRAD-Forêt,
45bis avenue de la Belle Gabrielle,
F-94736 Nogent-sur-Marne, France
RESUME
En relation avec une évaluation d'ensemble des ressources génétiques d'Acacia albida, on a étudié sa variation génétique et son système de reproduction, en utilisant des isozymes comme marqueurs génétiques.2 Les 19 populations étudiées ont montré un degré élevé de variation des isozymes à l'intérieur d'une même population et entre populations. Les principales différences ont été observées entre populations occidentales et orientales. Le degré d'allofécondation de 4 populations du Niger a été étudié; et s'est avéré plus élevé qu'on ne pouvait l'attendre d'une espèce entomophile, notamment si l'on considère le fait que la morphologie de l'inflorescence et la dynamique de la floraison sembleraient favoriser un degré élevé d'autofécondation. L'hypothèse est émise qu'Acacia albida posséderait un système de reproduction souple comportant un certain degré d'auto-incompatibilité.
INTRODUCTION
Acacia albida (syn. Faidherbia albida) est largement répandu en Afrique (Wickens, 1969), où il pousse dans des conditions variées de pluviométrie, de sols et d'altitude. Son importance comme ligneux à usages multiples en agroforesterie en Afrique est bien connue. L'arbre étant feuillé et fleurissant à la saison sèche, il fournit à cette période du fourrage tant par son feuillage que par ses gousses, ce qui est particulièrement important en fin de saison sèche, alors que les autres sources de fourrage se raréfient. Il a d'autre part une action améliorante sur la fertilité du sol, et fournit une large gamme d'autres produits utiles pour les communautés rurales.
En raison des menaces qui pèsent sur une partie de son ensemble génétique, de son importance dans le développement rural et des perspectives de domestication et d'amélioration génétique qu'il offre, A. albida a été dès 1974 classé comme essence prioritaire par le Groupe FAO d'experts des ressources génétiques forestières (FAO, 1974). C'est également l'une des essences faisant l'objet du projet FAO en cours sur les “Ressources génétiques de ligneux à usages multiples des zones sahélienne et nord-soudanienne d'Afrique” (de Framond, 1990). En dépit de ces activités, toutefois, on manque toujours d'information de base sur cette essence, notamment en ce qui concerne sa variation génétique et ses systèmes de reproduction.
La variation génétique peut être évaluée par des essais de provenances et de descendance, établis sur une gamme de stations. Après un travail de récolte systématique, complétant les récoltes effectuées dans le cadre du projet FAO mentionné ci-dessus et mené par les instituts de recherche forestière d'Afrique occidentale et le CTFT (maintenant CIRAD-Forêt), des essais de cette nature ont été mis en place en 1985 au Burkina Faso et au Zimbabwe. Au Burkina Faso, ils se situent à Gonsé (pluviométrie 800 mm) et à Dindéresso (pluviométrie 950 mm), et comprennent 19 populations d'Afrique occidentale (Burkina Faso, Mali, Niger, Sénégal), d'Afrique centrale (Cameroun) et d'Afrique orientale (Burundi, Ethiopie, Zimbabwe). Certaines de ces populations ont également fait l'objet d'essais au Zimbabwe. Des récoltes complémentaires sont en cours en Afrique orientale au titre d'un programme de l'Overseas Development Administration du Royaume-Uni (Fagg & Barnes, 1990). Des essais de terrain ont également été récemment mis en place dans d'autres pays. Bien que les résultats ne soient pas concluants, en partie en raison du mauvais choix d'une des stations du Burkina Faso où la mortalité a été élevée, il semble y avoir une nette différence de comportement entre les populations orientales et occidentales de l'espèce, qui se manifeste tant dans les essais du Burkina Faso (Billand, 1991) que dans ceux du Zimbabwe (Sniezko & Stewart, 1989).
Des essais de descendance ont été mis en place au Burkina Faso à partir de 1987 en vue d'estimer l'héritabilité de certains caractères importants de quelques provenances. Les premiers résultats de ces essais montrent une héritabilité de 0,26 pour la hauteur, à l'âge de 42 mois (Billand, 1991).
MATERIEL ET METHODES
On a à ce jour analysé les semences de 19 populations. Ces semences avaient été fournies par le CIRAD/CTFT. On a utilisé dans l'étude une graine par arbre, et on a pris une vingtaine d'arbres par population (Tableau 1).
Tableau 1. Origines des populations étudiées
| Pays | Population | Latitude | Longitude | Altitude | Pluviométrie |
| Burkina | Diou | 11°45'N | 02°56'W | 275 m | 800 mm |
| Faso | Dori | 14°02'N | 00°02'W | 275 m | 800 mm |
| Safané | 12°08'N | 03°13'W | 293 m | 800 mm | |
| Cameroun | Bibémi | 09°16'N | 13°50'E | 220 m | 875 mm |
| Guétalé | 10°57'N | 13°55'E | 450 m | 850 mm | |
| Makary | 12°34'N | 14°28'E | 286 m | 500 mm | |
| Mora | 11°00'N | 14°13'E | 400 m | 680 mm | |
| Moulvouday Zamay | 10°23'N | 14°50'E | 330 m | ||
| Mali | Barouéli | 13°05'N | 06°51'W | 250 m | 800 mm |
| Kémény | 12°58'N | 05°40'W | 270 m | 760 mm | |
| Kolongotomo | 13°49'N | 05°48'W | 280 m | 650 mm | |
| Samaye | 13°50'N | 04°45'W | 260 m | 570 mm | |
| Sarro | 13°42'N | 05°15'W | 250 m | 585 mm | |
| Niger | Bouza | 14°25'N | 06°07'E | 300 m | 690 mm |
| Kollo | 13°18'N | 02°21'E | 210 m | 593 mm | |
| Matameye | 13°25'N | 08°28'E | 450 m | 560 mm | |
| Tera | 14°00'N | 00°45'E | 240 m | 458 mm | |
| Zimbabwe | Mana pools | 15°45'S | 29°20'E | 360 m | 780 mm |
Pour la partie de l'étude concernant le flux de gènes à l'intérieur des populations, on a choisi les quatre populations du Niger. Six graines par arbre furent analysées, en utilisant l'électrophorèse sur des semis émergents âgés de 5 jours. Huit systèmes enzymatiques, représentant 10 loci sur 15 pour lesquels des procédures de coloration ont été décrites par Joly et al. (1992), ont été étudiés ici.
Pour chaque population on a calculé les paramètres suivants: nombre de loci polymorphes, nombre moyen d'allèles par locus, hétérozygotie observée, index de fixation (Tableau 2). La distribution de la variation entre populations et à l'intérieur des populations a été estimée selon Wright (1965). Une analyse en composantes principales a également été effectuée sur les fréquences observées d'allèles des populations.
Le flux de gènes à l'intérieur d'une population a été analysé suivant le modèle d'estimation multilocus du taux d'allofécondation de Ritland & Jain (1981), en utilisant un programme proposé par Ritland (1990).
Tableau 2. Caractéristiques génétiques des populations étudiées
| Hétérozygotie moyenne | ||||||
| Population | No. moyen l'allèles par locus | Pourcentage de loci poly morphes* | Comptage direct | Attendu** | Index de fixation | |
| 1. | DIOU (BF) | 2.8 | 80.0 | 0.298 | 0.371 | 0.357 |
| 2. | DORI (BF) | 3.3 | 90.0 | 0.330 | 0.444 | 0.227 |
| 3. | SAFANE (BF) | 2.5 | 80.0 | 0.228 | 0.293 | 0.392 |
| 4. | BIBEMI (C) | 2.9 | 80.0 | 0.365 | 0.437 | 0.175 |
| 5. | GUETALE (C) | 3.0 | 90.0 | 0.419 | 0.461 | 0.195 |
| 6. | MAKARY (C) | 2.6 | 90.0 | 0.200 | 0.343 | 0.417 |
| 7. | MORA (C) | 2.6 | 90.0 | 0.197 | 0.343 | 0.426 |
| 8. | MOUVOULDAY (C) | 3.3 | 90.0 | 0.360 | 0.446 | 0.193 |
| 9. | ZAMAY (C) | 3.3 | 90.0 | 0.274 | 0.377 | 0.165 |
| 10. | BAROUELI (M) | 2.9 | 90.0 | 0.267 | 0.415 | 0.091 |
| 11. | KEMENY (M) | 3.4 | 90.0 | 0.359 | 0.446 | 0.273 |
| 12. | KOLONGOTOMO (M) | 3.4 | 90.0 | 0.360 | 0.466 | 0.146 |
| 13. | SAMAYE (M) | 2.9 | 90.0 | 0.250 | 0.411 | 0.191 |
| 14. | SARRO (M) | 3.6 | 90.0 | 0.395 | 0.479 | 0.213 |
| 15. | BOUZA (N) | 3.0 | 90.0 | 0.340 | 0.398 | 0.345 |
| 16. | KOLLO (N) | 3.3 | 90.0 | 0.368 | 0.452 | 0.136 |
| 17. | MATAMEYE (N) | 3.3 | 90.0 | 0.325 | 0.405 | 0.257 |
| 18. | TERA (N) | 2.8 | 90.0 | 0.269 | 0.414 | 0.197 |
| 19. | MANA-POOLS (Z) | 2.4 | 50.0 | 0.229 | 0.265 | 0.222 |
** Estimation non biaisés (voir Nei, 1978)
Source: Joly, 1991
RESULTATS
Variation génétique à l'Intérieur des populations
Les résultats ont montré que les populations ont une variabilité élevée pour les loci examinés. Dans la plupart des cas, le pourcentage de loci polymorphes est de 90%. Seule la population de Mana Pools (Zimbabwe) montre un faible degré de variabilité (Tableau 2); c'est aussi la population qui présente le plus faible nombre d'allèles par locus. Le degré d'hétérozygotie, qui peut être considéré comme une mesure de la diversité allélique (Nei, 1977), est très élevé pour les populations d'Afrique occidentale (0,400) en comparaison de celles d'Afrique orientale (0,265).
Pour toutes les populations étudiées, les index de fixation moyens constatés indiquent un déficit d'hétérozygotes par rapport à une population panmictique. Certaines populations, telles que celles de Makary et de Mora au Cameroun, montrent un déficit très important (F.0,400), tandis que d'autres suivent un modèle attendu à la suite de panmixie.
Variation génétique entre populations
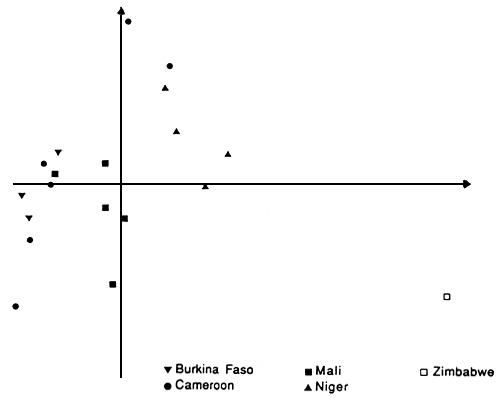
L'analyse en composantes principales (ACP) indique de grandes différences entre populations d'Afrique occidentale et d'Afrique orientale. Les deux premiers axes de l'ACP représentent 47% de la variation totale. Le premier axe oppose les populations occidentales à celles d'Afrique orientale. Le second axe traduit des différences entre les populations occidentales; deux populations du Cameroun (Bibémi et Guétalé) ainsi que des populations du Niger semblent être différentes du reste (Figure 1). Le dendrogramme obtenu avec la distance génétique absolue entre populations vient à l'appui de ces constatations (Joly et al., 1992).
Figure 1. Projection sur le plan (1,2) de l'analyse en composantes principales des populations étudiées
Système de reproduction
Le taux d'allofécondation (tm) estimé pour les quatre populations étudiées varie entre 0,893 et 0,996 (Joly & Aygalent, 1992); il est supérieur à la valeur attendue (tm < 0,90) pour une espèce entomophile (Surles et al., 1990). La durée de la floraison chez Acacia albida est de 10 à 14 semaines (Zeh-Nlo & Joly, sous presse); le nombre de fleurs hermaphrodites par inflorescence est élevé (une centaine), et ces fleurs s'ouvrent progressivement (Tybirk & Jørgensen, sous presse; Zeh-Nlo, données non publiées). Pour cette raison, on s'attendrait à un taux élevé d'autopollinisation chez Acacia albida. Les résultats présentés dans cet article semblent indiquer qu'A. albida possède un système d'auto-incompatibilité, qui semble fonctionner avec une intensité variable selon les individus, et ceux-ci présentent en conséquence une grande variation dans leur degré d'allofécondation (Joly & Aygalant, 1992).
CONCLUSION
Cette étude indique qu'Acacia albida est une espèce génétiquement variable, qui présente un degré élevé de variation génétique par comparaison avec des acacias australiens qui ont été soumis à des études analogues. D'autres résultats sont encore attendus sur les populations orientales, mais on peut dire que ces premiers résultats sont comparables aux premières constatations faites sur A. albida, faisant apparaître des différences importantes entre populations occidentales et orientales de l'espèce en ce qui concerne les caractères de croissance. Les résultats indiquent également qu'A. albida est auto-incompatible à des degrés divers.
D'autres études de plus grande ampleur devront encore être effectuées, en utilisant des marqueurs génétiques aussi bien que des essais de terrain, pour avoir un tableau général des modèles de variation génétique chez A. albida. Des croisements dirigés devront être effectues à l'appui de ces études, avec du pollen propre et du pollen étranger, pour clarifier les aspects relatifs aux systèmes de reproduction (Joly, 1991).
Les présents résultats peuvent être considérés comme un premier pas vers une meilleure connaissance de cette espèce, et ils devront être pris en compte dans l'élaboration de programmes d'amélioration et de conservation.
REMERCIEMENTS
Cette étude a été financée au titre du programme de recherche de la CEE STD2 n° TS2*0207 -F(EDB).
REFERENCES
BILLAND, A. 1991 Variabilité génétique de Faidherbia albida en essais comparatifs de descendances au Burkina Faso. Actes de l'atelier international sur Faidherbia albida en Afrique occidentale tenu à Niamey, 22–26 avril 1991 (sous presse).
BILLAND, A. & DE FRAMOND, H. 1990 Variabilité génétique d'Acacia albida (synonyme Faidherbia albida) en essais de provenances au Burkina Faso. Actes del'atelier sur la Physiologie des arbres et arbustes en zones arides et semi-arides, Nancy, Avril 1990.
DANTHU, P. & PRAT, D. 1991 Study of the genetic variability by means of isoenzymes in Faidherbia albida; preliminary results. In Biochemical markers in the population genetics of forest trees, S. Fineschi M.E. Malvoti, F Cannata and H.H. Hattemer editors. SPB Academic Publishing by The Hague.
DE FRAMOND, H. 1990 Valorisation des ressources génétiques des ligneux à usages multiples en Afrique soudano-sahélienne. IRGF 18: 21–27.
FAO 1974 Troisième Session du Groupe FAO d'Experts des Ressources Génétiques Forestières. Rome, Italie, 6–10 mai 1974. 90 p.
FAGG, C.W. and BARNES, R.D. Africa acacias: study and acquisition of the genetic resources. ODA research scheme R.4348 Final Report, Oxford Forestry Institute, 170 pp.
JOLY, H.I. 1991 Population genetics of Acacia albida (syn. Faidherbia albida). Bull.Int. Group Study of Mimosoideae, 19:86–95.
JOLY, H.I. 1991 Acacia albida: aspects génétiques. Actes de l'atelier international surFaidherbia albida en Afriques occidentale. Niamey, 22–26/4–91 (sous presse).
JOLY, H.I. ZEH-NLO, M., DANTHU, P. & AYGALENT, C. 1992 Population genetics of an African Acacia: Acacia albida. Genetic diversity of populations from West Africa. Aust. J. Bot. 40:59–73.
JOLY, H.I. & AYGALENT, C. 1992 Breeding system of Acacia albida (syn. Faidherbia albida). Preliminary results. In “Population genetics and gene conservation in forest trees”, IUFRO symposium, Carcans Maubuisson, 24–28 August 1992.
NEI, M.F. 1977 Statistics and analysis of gene diversity in subdivided populations. Ann. Human Genet. 41:225–233.
RITLAND, K. 1990 A series of FORTRAN computer programs for estimating plant mating systems. J. Heredity, 81:235–237.
RITLAND, K. & JAIN, S. 1981 A model for the estimation of outcrossing rate and gene frequencies using an independent loci. Heredity, 47:35–52.
SNIEZKO, R.A. & STEWART, H.T.L. 1989 Range-wide provenance variation in growth and nutrition of Acacia albida seedlings propagated in Zimbabwe. Forest Ecology and Management 27:179–197.
SURLES, S., HAMRICK, J. & BONGARTEN, B. 1990 Mating systems in open pollinated families of black locust (Robinia pseudoacacia). Silvae Genet. 39:35–40.
TYBIRK, K. & JORGENSEN, A. 1991 Floral biology and pollination of some African acacias and Faidherbia albida. In the proceedings of the A.E.T.A.T. meeting held in Malawi 1991 (in press).
WICKENS, G.E. 1969 A study of Acacia albida Del. (Mimosoideae). Kew Bull. 23:181–202
WRIGHT, S. 1965 The interpretation of population structure by F-statistics with special regard to systems of mating. Evolution 19:395–420.
ZEH-NLO, M. & JOLY, H. 1991 Observations préliminaires sur la phénologie d'Acacia albida Etude d'une population du nord Cameroun. Actes de l'atelier international sur Faidherbia albida en Afrique occidentale à Niamey, 22–26/4–91 (sous presse).
![]()
![]()
![]()