![]()
![]()
![]()
H.I. Joly1
Laboratorio INRA-ENGREF de Ciencias Forestales
Unidad de Genética de poblaciones de árboles forestales
asociado con el CIRAD-Forêt, Francia
Dirección actual: CIRAD-Forêt
45 bis avenue de la Belle Gabrielle,
94 736 Nogent-sur-Marne, Francia
RESUMEN
A propósito de una evaluación general de los recursos genéticos de Faidherbia albida, se estudió su variación genética y su sistema de mejoramiento genético empleando isoenzimas como indicadores genéticos2. Las 19 poblaciones estudiadas mostraron un alto nivel de variación en isoenzimas dentro de las poblaciones y entre ellas. Las diferencias principales se daban entre las poblaciones occidentales y las orientales. Se estudió el nivel de cruzamiento lejano de 4 poblaciones procedentes de Níger y se encontró que era extraordinariamente alto para una especie entomógama, considerando sobre todo el hecho de que la morfología de la influorescencia y la dinámica de la floración parecen favorecer un alto grado de autopolinización. Ello sugiere que la Faidherbia albida posee un sistema flexible de cruzamiento que incluye una cierta incompatibilidad para la autopolinización.
INTRODUCCION
La Faidherbia albida (sinónimo Acacia albida) está extensamente distribuida por Africa (Wickens 1969) donde se desarrolla en condiciones diversas de precipitaciones, suelos y altitudes. Es bien conocida su importancia como árbol de finalidad múltiple en la agrosilvicultura de Africa. Como la especie mantiene las hojas y florece durante la estación seca, se dispone de hojas y legumbres como forraje, lo que es especialmente importante al final de la estación seca, cuando escasean otras fuentes de forraje. El árbol favorece también a la fertilidad del suelo y produce una amplia variedad de otros productos que utilizan las comunidades rurales.
Debido a las amenazas a ciertas partes de su caudal genético, a su importancia para el desarrollo rural y al campo que existe para su tratamiento y mejora, la F. albida ha sido incluida desde 1974 (FAO 1974) como especie prioritaria por el Cuadro de Expertos de la FAO en Recursos Genéticos Forestales, (FAO 1974). Es también una de las especies incluidas en el Proyecto en ejecución, de la FAO, sobre Recursos Genéticos de Especies Leñosas de Finalidad Múltiple, de las zonas africanas del Sahel y del Norte de Sudán (de Framond 1990). Sin embargo, a pesar de estas actividades, falta todavía información básica sobre esta especie, particularmente de información relacionada con la variación genética y con los sistemas de mejoramiento genético.
La variación genética se puede evaluar mediante ensayos de procedencias y progenies, establecidos en diversas estaciones. Después de un trabajo sistemático de recolección, para complementar las recolecciones realizadas dentro del marco del proyecto de la FAO antes mencionado, llevado a cabo por institutos nacionales de investigación de Africa occidental y del CTFT (ahora CIRAD-Forêt), se establecieron tales ensayos en Burkina Faso y en Zimbabwe, en 1985. En Burkina Faso, se establecieron los ensayos en Gonsé (precipitaciones de 800 mm.) y en Dindéresso (precipitaciones de 950 mm.), empleando 19 poblaciones procedentes de Africa Occidental (Burkina Faso, Mali, Níger, Senegal), Africa Central (Camerún) y Africa Oriental (Burundi, Etiopía, Zimbabwe). Algunas de las mismas poblaciones se ensayaron también en Zimbabwe. Se están realizando recolecciones adicionales en Africa Oriental bajo el Programa de la Administración para el Desarrollo Internacional (Overseas Development Administration) (Reino Unido) (Fagg & Barnes 1990). Recientemente se han establecido también ensayos de campo en otros países. Aunque los resultados no fueron concluyentes, debido en parte al carácter inapropiado de una de las estaciones de Burkina Faso donde la mortalidad fue elevada, parece existir una clara diferencia entre el comportamiento de las poblaciones orientales y las occidentales de la especie, que resultó evidente tanto en los ensayos de Burkina Faso (Billand, 1991) como en los de Zimbabwe (Sniezko & Stewart 1989).
En Burkina Faso se han establecido ensayos de progenie desde 1987, a fin de determinar el carácter hereditario de ciertos rasgos importantes de algunas procedencias. Los primeros resultados de estos ensayos han demostrado una transmisión por herencia de 0,26 en cuanto a la altura, a los 42 meses de edad (Billand, 1991).
Los ensayos de campo antes descritos permitirán avanzar en la determinación de la variación genética y el comportamiento hereditario de la Faidherbia albida. Sin embargo, los estudios de campo sobre la diversidad genética se pueden complementar mediante estudios de índices genéticos, permitiendo también esta última metodología la estimación del flujo de genes dentro de las poblaciones y entre ellas. El presente documento describe el trabajo realizado sobre genética de poblaciones de F.albida, en el Laboratorio de INRA-ENGREF de Nancy, mediante el análisis de las semillas utilizando isoenzimas como indicadores genéticos.
MATERIAL Y METODOS
Hasta ahora se han analizado semillas procedentes de 19 poblaciones. Las semillas fueron proporcionadas por CIRAD-CTFT. En este estudio se empleó una semilla por árbol y alrededor de 20 árboles por población (véase el Cuadro 1).
Cuadro 1: Orígenes de las poblaciones estudiadas
| País | Población | Latitud | Longitud | Altitud | Precipitación |
|---|---|---|---|---|---|
| Burkina Faso | Diou | 11°45'N | 02°56'W | 275 m. | 800 mm. |
| Dori | 14°02'N | 00°02'W | 275 m. | 800 mm. | |
| Safané | 12°08'N | 03°13'W | 293 m. | 800 mm. | |
| Camerún | Bibémi | 09°16'N | 13°50'E | 220 m. | 875 mm. |
| Guétalé | 10°57'N | 13°55'E | 450 m. | 850 mm. | |
| Makary | 12°34'N | 14°28'E | 286 m. | 500 mm. | |
| Mora | 11°00'N | 14°13'E | 400 m. | 680 mm. | |
| Moulvouday Zamay | 10°23'N | 14°50'E | 330 m. | ||
| Mali | Barouéli | 13°05'N | 06°51'W | 250 m. | 800 mm. |
| Kémény | 12°58'N | 05°40'W | 270 m. | 760 mm. | |
| Kolongotomo | 13°49'N | 05°48'W | 280 m. | 650 mm. | |
| Samaye | 13°50'N | 04°45'W | 260 m. | 570 mm. | |
| Sarro | 13°42'N | 05°15'W | 250 m. | 585 mm. | |
| Níger | Bouza | 14°25'N | 06°07'E | 300 m. | 690 mm. |
| Kollo | 13°18'N | 02°21'E | 210 m. | 593 mm. | |
| Matameye | 13°25'N | 08°28'E | 450 m. | 560 mm. | |
| Tera | 14°00'N | 00°45'E | 240 m. | 458 mm. | |
| Zimbabwe | Mana Pools | 15°45'S | 29°20'E | 360 m. | 780 mm. |
(De Joly 1991)
Para el estudio parcial referente al flujo de genes dentro de las poblaciones, se eligieron las 4 poblaciones disponibles de Níger. Se analizaron 6 semillas por árbol, empleando la electroforesis en brinzales emergentes de 5 días. En el presente estudio se investigaron 8 sistemas enzimáticos, que representaban 10 “loci” de los 15 para los que Joly et al (1992) han descrito procedimientos de teñido.
Para cada población se calcularon los siguientes parámetros genéticos: número de loci polimórficos, número medio de alelos por “locus”, heterozigosidad observada, heterozigosidad esperada e índices de fijación (véase el Cuadro 2). La distribución de la variación entre poblaciones y dentro de ellas se calculó siguiendo a Wright (1965). Se realizó también un análisis de componentes principales sobre las frecuencias observadas de alelos en las poblaciones.
El flujo de genes dentro de una población se analizó siguiendo el modelo de estimación de la tasa de cruzamiento lejano de multilocus de Ritland & Jain (1981), utilizando un programa proporcionado por Ritland (1990).
Cuadro 2: Características genéticas de las poblaciones estudiadas
| Media de heterozigosidad | ||||||
| Población | Media de alelos por locus | Porcentaje de loci polimórficos* | Conteo directo | Esperado** | Indice de fijación | |
|---|---|---|---|---|---|---|
| 1. | DIOU (BF) | 2,8 | 80,0 | 0,298 | 0,371 | 0,357 |
| 2. | DORI (BF) | 3,3 | 90,0 | 0,330 | 0,444 | 0,227 |
| 3. | SAFANE (BF) | 2,5 | 80,0 | 0,228 | 0,293 | 0,392 |
| 4. | BIBEMI (C) | 2,9 | 80,0 | 0,365 | 0,437 | 0,175 |
| 5. | GUETALE (C) | 3,0 | 90,0 | 0,419 | 0,461 | 0,195 |
| 6. | MAKARY (C) | 2,6 | 90,0 | 0,200 | 0,343 | 0,417 |
| 7. | MORA (C) | 2,6 | 90,0 | 0,197 | 0,343 | 0,426 |
| 8. | MOUVOULDAY (C) | 3,3 | 90,0 | 0,360 | 0,446 | 0,193 |
| 9. | ZAMAY (C) | 3,3 | 90,0 | 0,274 | 0,377 | 0,165 |
| 10. | BAROUELI (M) | 2,9 | 90,0 | 0,267 | 0,415 | 0,091 |
| 11. | KEMENY (M) | 3,4 | 90,0 | 0,359 | 0,446 | 0,273 |
| 12. | KOLONGOTOMO (M) | 3,4 | 90,0 | 0,360 | 0,466 | 0,146 |
| 13. | SAMAYE (M) | 2,9 | 90,0 | 0,250 | 0,411 | 0,191 |
| 14. | SARRO (M) | 3,6 | 90,0 | 0,395 | 0,479 | 0,213 |
| 15. | BOUZA (N) | 3,0 | 90,0 | 0,340 | 0,398 | 0,345 |
| 16. | KOLLO (N) | 3,3 | 90,0 | 0,368 | 0,452 | 0,136 |
| 17. | MATAMEYE (N) | 3,3 | 90,0 | 0,325 | 0,405 | 0,257 |
| 18. | TERA (N) | 2,8 | 90,0 | 0,269 | 0,414 | 0,197 |
| 19. | MANA POOLS (Z) | 2,4 | 50,0 | 0,229 | 0,265 | 0,222 |
* Un locus se considera polimórfico si la frecuencia del alelo más corriente no excede de 0,95.
** Estimación imparcial (véase Nei, 1978)
(Tomado de Joly, 1991)
RESULTADOS
Variación genética dentro de las poblaciones
Los resultados demostraron que las poblaciones eran muy variables para los loci examinados. En la mayoría de los casos el porcentaje de loci polimórficos era del 90%; sólo la población de Mana Pools, Zimbabwe, presentó un bajo nivel de diversidad (véase el Cuadro 2); ésta fue también la población con el menor número de alelos por locus. El nivel de heterozigosidad, que puede considerase como una medida de la diversidad alélica (Nei 1977), fue muy elevado para las poblaciones de Africa Occidental (0,400) en comparación con las de Africa Oriental (0,265).
Para todas las poblaciones estudiadas, los índices medios de fijación hallados indicaron un déficit en heterozigotos en comparación con una población panmíctica. Algunas poblaciones como las de Makary y Mora de Camerún, mostraron un déficit muy grande (F. 0,400), mientras que otras siguieron el modelo esperado después de la panmixis.
Variación genética entre poblaciones
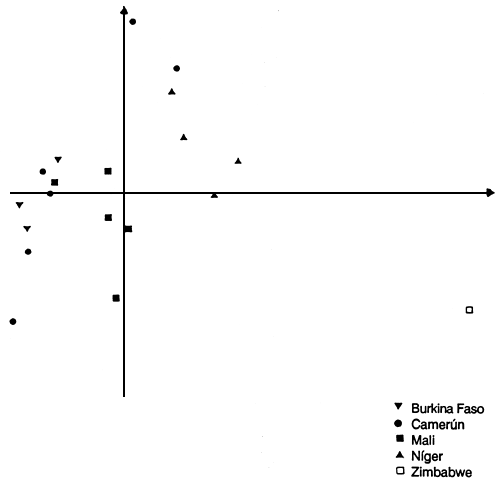
El análisis de componentes principales (ACP) indicó que existían grandes diferencias entre las poblaciones procedentes de Africa Occidental y las de Africa Oriental. Los dos primeros axis del ACP representaron el 47% de la variación total. El primer axis mostraba la diferencia entre las poblaciones occidentales y la oriental. El segundo axis incluía las diferencias entre las poblaciones occidentales; las dos poblaciones procedentes de Camerún (Bibémi y Guétale), junto con las poblaciones procedentes de Níger parecían ser diferentes del resto (Figura 1). El dendrograma obtenido con la distancia genética absoluta entre las poblaciones sirvieron de apoyo a estas conclusiones (Joly et al 1992).
Figura 1: Proyección de la población estudiada sobre el plano (1,2) del análisis de componentes principales
Sistema de mejoramiento genético
La tasa estimada de crecimiento lejano (tm) para las cuatro poblaciones estudiadas variaba de 0,893 a 0,996 (Joly y Aygalent 1992); éste es más elevado que el valor esperado para una especie estomógama (tm<0,90) (Surles et al 1990). La duración de la floración de la F.albida es de 10 a 14 semanas (Zeh-Nlo y Joly, en prensa); el número de flores hermafroditas de una influorescencia es grande (alrededor de 100) y estas flores se abren progresivamente (Tybirk y Jorgensen, en prensa, ZehNlo, datos sin publicar). En vista de ésto, podría esperarse una gran cantidad de autopolinización en la F.albida. Los resultados presentados en este documento sugieren que la F.albida posee un sistema de auto-incompatibilidad, que parece actuar con intensidad variable para árboles individuales, lo que consecuentemente varía muchísimo su nivel de cruzamiento lejano (Joly y Aygalant 1992).
CONCLUSION
Este estudio indica que la F.albida es una especie genéticamente variable, con un alto grado de variación genética en comparación con las acacias australianas que han sido objeto de estudios similares. Aunque se están obteniendo todavía más resultados sobre las poblaciones orientales, estos primeros resultados son comparables a las primeras conclusiones relacionadas con la F.albida, que señalan grandes diferencias entre las poblaciones occidentales y orientales de la especie, respecto a los caracteres de crecimiento. Los resultados indican también que la F.albida es, con diversos niveles, auto-incompatible.
Habrá que realizar nuevos y más amplios estudios, utilizando indicadores genéticos y ensayos de campo, para obtener una imagen general de los modelos de variación genética de la F.albida. Deben realizarse cruzamientos controlados en apoyo de tales estudios, incluyendo polen propio y externo, para aclarar aspectos relativos a los sistemas de mejora genética (Joly 1991).
Los presentes resultados pueden considerarse como un primer paso para un mejor conocimiento de la especie, debiendo considerarse debidamente en el desarrollo de programas de mejora y conservación.
AGRADECIMIENTO
Este estudio ha sido financiado dentro del programa de investigación de la CEE, STD2 no TS2*0207 -F(EDB).
REFERENCIAS
BILLAND, A. 1991 Variabilité génétique de Faidherbia albida en essais comparatifs de descendances au Burkina Faso. Actas del seminario internacional sobre la Faidherbia albida en Africa Occidental celebrado en Niamey, 22–26 Abril, 1991 (en prensa).
BILLAND, A. & DE FRAMOND, H. 1990 Variabilité génétique d'Acacia albida (synonyme Faidherbia albida) en essais de provenances au Burkina Faso. Actas del seminario “Physiologie des arbres et arbustes en zones arides et semi-arides”, Abril 1990.
DANTHU, P. & PRAT, D. 1991 Study of the genetic variability by means of isoenzymes in Faiherbia albida; preliminary results. In Biochemical markers in the population genetics of forest trees, S. Fineschi M.E. Malvoti, F. Cannata and H.H. Hattemer editors. SPB Academic Publishing by The Hague.
DE FRAMOND, H. 1990 Desarrollo de recursos genéticos de árboles de finalidad múltiple en las zonas africanas del Sudán y el Sahel. Información sobre recursos genéticos forestales. No 18. pp. 21–27. Trees in Sudano-Sahelian Africa. Forest Genetic Resources Information 18:21–27.
FAO 1974 Tercera Reunión del Cuadro de Expertos de la FAO en Recursos Genéticos Forestales. Celebrada en Roma, Italia, 6–10 Mayo 1974. 90 p.
FAGG, C.W. and BARNES, R.D. Africa acacias: study and acquisition of the genetic resources. ODA research scheme R.4348 Final Report, Oxford Forestry Institute, 170 pp.
JOLY, H.I. 1991 Population genetics of Acacia albida (syn. Faidherbia albida). Bull. Int. Group Study of Mimosoideae, 19:86–95.
JOLY, H.I. 1991 Acacia albida: genetic aspects. Actas del seminario internacional sobre la Faidherbia albida en Africa Occidental, celebrado en Niamey 22–26 Abril 1991 (en prensa).
JOLY, H.I. ZEH-NLO, M., DANTHU, P. & AYGALENT, C. 1992 Population genetics of an African Acacia: Acacia albida. Genetic diversity of populations from West Africa. Aust. J. Bot. 40:59–73.
JOLY, H.I. & AYGALENT, C. 1992 Breeding system of Acacia albida (syn. Faidherbia albida). Preliminary results. In “Population genetics and gene conservation in forest trees”, IUFRO symposium, Carcans Maubuisson, 24–28 Agosto 1992.
NEI, M.F. 1977 Statistics and analysis of gene diversity in subdivided populations. Ann. Human Genet. 41:225–233.
RITLAND, K. 1990 A series of FORTRAN computer programs for estimating plant mating systems. J. Heredity, 81:235–237.
RITLAND, K. & JAIN, S. 1981 A model por the estimation of outcrossing rate and gene frequencies using an independent loci. Heredity, 47:35–52.
SNIEZKO, R.A. & STEWART, H.T.L. 1989 Range-wide provenance variation in growth and nutrition of Acacia albida seedlings propagated in Zimbabwe. Forest Ecology and Management 27:179–197.
SURLES, S.E., HAMRICK, J.L. & BONGARTEN, B.C. 1990 Mating systems in open-pollinated families of black locust (Robinia pseudoacacia). Silvae Genet. 39:35–40.
TYBIRK, K. & JORGENSEN, A. 1991 Floral biology and pollination of some African acacias and Faidherbia albida. Actas de la reunión de la A.E.T.A.T. celebrada en Malawi 1991 (en prensa).
WICKENS, G.E. 1969 A study of Acacia albida Del. (Mimosoideae. Kew Bull. 23:181–202.
WRIGHT, S. 1965 The interpretation of population structure by F-statistics with special regard to systems of mating. Evolution 19:395–420.
ZEH-NLO, M. $ JOLY, H.I. 1991 Observations préliminaires sur la phénologie d'Acacia albida: Etude d'une population du nord Cameroun. Actas del seminario internacional sobre la Faidherbia albida en Africa Occidental, celebrado en Niamey 22–26 Abril 1991 (en prensa).
![]()
![]()
![]()