![]()
![]()
![]()
Por J. P. PERRY, Jr.,
Oficial Ejecutivo del Programa Agrícola del Instituto Rockefeller en México
Las informaciones disponibles revelan que la muerte de la madera de pino en extensas zonas de México Central fué atribuida por primera vez en 1903, al escarabajo taladrador de cortezas. En dicho año, el Dr. Sylvio J. Bonansea fué designado por el Gobierno de México para investigar la causa de extinción de los pinares en los Estados mexicanos de Morelos y Michoacán, a cuyos habitantes preocupaba la desaparición de grandes cantidades de pinos. El Dr. Bonansea descubrió que el escarabajo taladrador de cortezas era la causa principal. En su informe titulado «La Plaga de Ocotes», que no se publicó hasta 1921, se describen los aspectos de los árboles muertos así como los insectos y su labor. También incluye muchas observaciones sobre las características generales de la plaga y sugiere varias medidas de control.
El Dr. Bonansea envió ejemplares de insectos recogidos en estas zonas al Dr. A.D. Hopkins, de la Oficina de Entomología de la Secretaría de Agricultura de los Estados Unidos de América. Sirviéndose de este material, el Dr. Hopkins describió el Dendroctonus mexicanus Hopk. y el Ips bonanseai Hopk. como especies nuevas. En su publicación, titulada «Genus Dendroctonus», Hopkins afirma que el Dendroctonus mexicanus «ataca, evidentemente, al Pinus teocotl Schl. y Cham.; al P. leiophylla Sch. y Cham., y al P. ayacahuite Ehr.» Se indica que en Michoacán se dan otras especies, como el Dendroctonus parallelocollis Hopk., pero no se mencionan los árboles hospederos. Una publicación posterior, de Donald De León, enumera cierto número de especies de escarabajos taladradores de corteza, que atacan al pino en México, con descripciones y referencias sobre los árboles hospederos. En la literatura de que se dispone hay muy poca información sobre la vida y hábito de estos insectos, su relación con los árboles hospederos o las causas de los violentos ataques que caracteriza a las infestaciones.
En la actualidad, existe una infestación muy epidémica de escarabajos taladradores en los rodales de Pinus leiophylla de los alrededores del Valle de México, infestación que, en todas sus características, se parece a los focos de hace medio siglo, descritos por el Dr. Bonansea.1 Recientes observaciones del autor indican que en México Central se han registrado focos diseminados de escarabajos taladradores a intervalos periódicos y en el curso de un largo período, y que la transición del tapiz forestal en terreno de aprovechamiento agrícola se acorta a veces por los efectos de las infestaciones del escarabajo taladrador.
[1 Actualmente está trabajando en México un entomólogo de la FAO J. M. Miller, que fué enviado a, ese país en virtud del Programa Ampliado de Asistencia Técnica. Otros dos técnicos de la FAO, G. Becker y F. Schwerdtfeger, han hecho también investigaciones acerca de otras plagas análogas de escarabajos de corteza en Guatemala.]
Este artículo tiene por objeto registrar los resultados de estudios realizados por el autor durante los años 1949 y 1950 en México. En el transcurso de excursiones intermitentes al campo, desde la capital de México, ha sido posible llegar al conocimiento de los hábitos de tres especies de escarabajos taladradores, sus relaciones recíprocas, la forma en que atacan y matan a los árboles, y la extensión en que pueden destruir ciertos rodales de pinos.
Zonas y Carácter de los Rodales de Pinos Afectados por las Recientes Epidemias.
Las zonas infestadas que ha estudiado el autor se encuentran cerca de las ciudades de Tlalmanalco, San Rafael y Amecameca, en el Estado de México. El Dr. Sylvio Bonansea se refiere a grandes infestaciones de D. mexicanus en esta misma localidad, e, indudablemente, en toda la zona ha habido plagas intermitentes durante los últimos 45 años. A juzgar por la extensión de las actuales zonas destruidas por el escarabajo taladrador, las pérdidas de madera en dicha localidad durante estos años deben haber sido enormes.
Las informaciones obtenidas de los moradores de la région, fechan el comienzo de la epidemia actual hacia el año 1947, y los peones interrogados coincidieron unánimemente en localizar el primer foco de la plaga en las laderas de un montículo a pocos kilómetros al oeste de la ciudad de Amecameca. El autor ha observado las últimas fases de esta destrucción, y ha observado que el bosque se componía de un rodal regular de Pinus leiophylla, 2 con un diámetro medio de 10 pulgs. (25 cms.) a la altura del pecho, y con una capa baja de chaparros (Quercus sp.) desperdigados. En los terrenos orientados al norte y al nordeste, los cedros (Cupressus benthamii Endl.) formaban también una pequeña parte de la capa alta. Con excepción de unos cuantos árboles del borde sudoccidental, este pinar ha desaparecido totalmente, no quedando más que chaparros y cedros. La superficie destruida se calcula en 4000 acres (1.600 Ha.). El D. mexicanus3 parece haberse extendido en 1949 desde la zona infestada por toda la campiña circundante atacando a muchos pequeños rodales aislados de pinos en los alrededores de Tlalmanalco. Fué precisamente sobre cierto número de estos rodales que el autor pudo seguir con bastante detalle el avance y desarrollo del D. mexicanus.
[2 Los ejemplares de árboles mencionados en este articulo están registrados en el herbario del autor.
3 Ejemplares duplicados de los insectos citados en este artículo están registrados en cl Musco Nacional de los Estados Unidos de América.]
Sin embargo, la dispersión de este insecto no acababa en estos rodales aislados. En 1950, se observó por primera vez un foco repentino, de una extensión de 5 acres (2 Ha.), aproximadamente, en las colinas al pie de la cordillera de Sierra Nevada cerca de la ciudad de San Rafael, a unas 5 millas (8 Kms.) de Amecameca. Este foco se está propagando ahora hasta los pinares de las laderas de esta cordillera, y sólo cabe calcular a la ligera el número de hectáreas de bosque que quedarán destruidas. El pino predominante en las laderas bajas de esta zona es el Pinus leiophylla, entremezclado con el P. montezumae y el P. montezumae var. macrocarpa Martínez. En los puntos más altos destacan el P. montezumae y el P. montezumae var. macrocarpa Martínez y comienza a aparecer el abeto (Abies religiosa), que aumenta en cantidad con la altura. A medida que el bosque varía gradualmente con la elevación, pasando del P. leiophylla predominante al P. montezumae var. macrocarpa - A. religiosa, la plaga va perdiendo aparentemente su fuerza y vigor y desaparece poco a poco.
 |
|
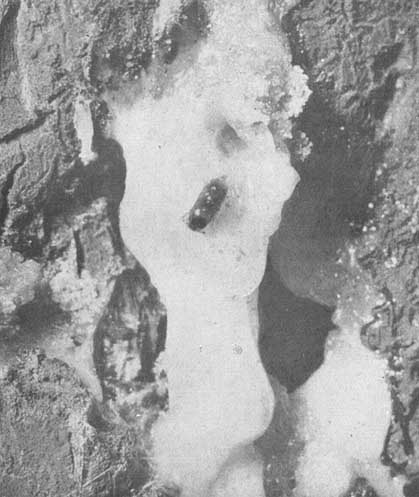
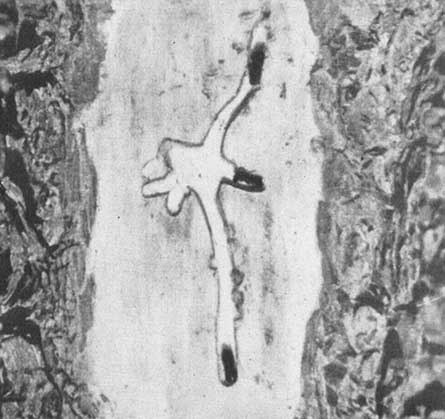
A la izquierda: un macho de Dendroctonus mexicanas perforando el chorro de resina para mantener abiertas la galería y la entrada. Arriba: una larva plenamente desarrollada y un joven adulto en las celdas de la corteza exterior del Pinus leiophylla.
Historia del Dendroctonus mexicanas
El D. mexicanus es el escarabajo taladrador principalmente responsable de las ingentes pérdidas en estas zonas. Los hábitos de esta especie no han sido descritos ampliamente en publicaciones anteriores, por lo que se tratan aquí con algún detalle. Hopkins describió en 1906 los adultos de esta especie, afirmando que miden un promedio de 3,8 mm. de longitud y que son muy parecidos al D. frontalis, aunque algo más pequeños. Los adultos atacan al árbol en grupos, pero cada entrada separada a la corteza representa el trabajo de un par de escarabajos, porque, por lo general, la hembra es en realidad la que hace la perforación de la entrada de la galería. Desde la entrada, excava una galería de corta distancia en dirección vertical ascendente, que permite que la resina fluya libremente por el orificio. Durante esta fase, la actividad del macho se limita a empujar a la resina para que salga de la galería. Cuando cesa el flujo de resina, hacen una curva cerrada en la galería y la prolongan hacia abajo y alrededor del tronco, en espiral, adoptando la forma de una «S» alargada. El autor ha registrado galerías de 13 a 16 pulgs. (33 a 41 cms.) de longitud, que los adultos continuaban alargando activamente y donde seguían desovando.
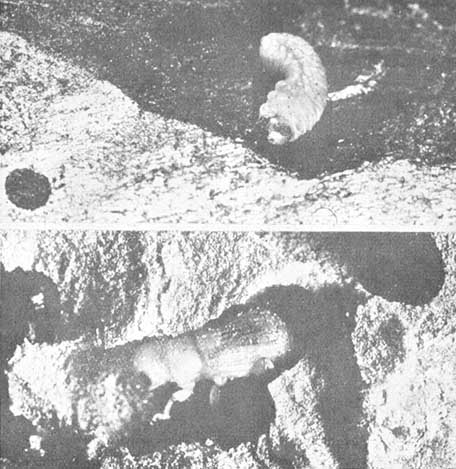
El desove tiene lugar del siguiente modo: la hembra excava primero un pequeño nicho en la pared de la galería, donde deposita el huevo, al que envuelve con excremento. Los huevos se depositan en nichos alternos, a ambos lados de la galería. De varios recuentos de huevos se deduce que la hembra deposita un promedio de seis por cada pulgada (2,5 cms.) de galería. El macho, que trabaja aproximadamente a 2 pulgs. (5 cms.) detrás de la hembra, va consolidando las paredes de la galería con excremento. Al romperse el huevo, las minúsculas larvas se abren camino perforando hasta el líber, en ángulo recto con la galería principal, y a medida que crecen, avanzan gradualmente alimentándose hacia la corteza exterior seca, alcanzando su máximo crecimiento al llegar a esta última. Al penetrar en la corteza cesa toda alimentación y construyen celdas, donde las larvas se transforman en crisálidas, luego en adultos jóvenes y, por último, en nuevos adultos. Estos se abren camino directamente desde la celda de la crisálida y a través de la corteza, emergen completamente desarrollados y en condiciones de iniciar inmediatamente un nuevo ataque contra los pinos vivos.
Forma característica de galería construída por escarabajos adultos en el Pinus leiophylla.
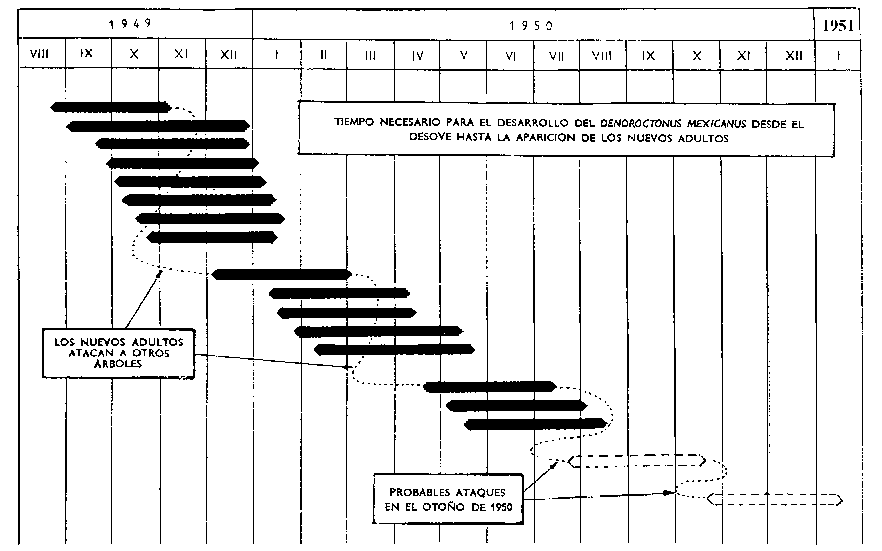
En el cuadro siguiente figuran los datos sobre el promedio de días necesarios para el desarrollo del D. mexicanus en el Pinus leiophylla :
CUADRO 1. - NUMERO DE DÍAS QUE SE REQUIEREN PARA LA TRANSFORMACIÓN DEL HUEVO DEL D. mexicanus EN INSECTO A ADULTO EN EL Pinus leiophylla.
|
N.° de la zona * |
N.° de árboles en que se siguió la desarrollo de la cría |
Promedio de los días necesarios para la evolución del huevo en adulto |
|
4 |
36 |
79,9 |
|
5 |
2 |
76,0 |
|
1 |
19 |
78,6 |
|
3 |
57 |
79,4 |
* Los números de zonas corresponden a rodales seleccionados que representan condiciones que varían ligeramente en elevación y orientación.
El Potencial Biótico. De los datos relativos al período de desarrollo se deduce que cada año pueden darse cuatro generaciones de insectos. Esto dota al insecto de un enorme potencial biótico - particularmente porque durante su ciclo vital no se ha observado ningún auténtico período de letargo. Incluso durante los meses más fríos del año, las temperaturas en el centro del día son lo suficientemente elevadas para asegurar cierta alimentación y el desarrollo de las larvas. El autor ha registrado también numerosos ataques eficaces durante esos meses.
En estos estudios no se dispuso de tiempo para deber. minar si después de construidas las galerías de desove en el primer árbol atacado, los adultos padres emergían otra vez, en cualquier proporción, para acometer de nuevo a otros árboles. Este hábito ha sido estudiado en una especie muy semejante de California, el D. brevicomis, observándose que, a veces, reaparecía hasta un cincuenta por ciento de adultos padres para iniciar nuevos ataques. Sin embargo, las observaciones de las galerías de desove construidas por el D. mexicanus indican que la mayoría de los adultos padres muere cerca del final de aquellas. La reaparición de los adultos padres no es, por lo visto, un factor importante en el potencial biótico de este escarabajo taladrador de cortezas.
El Dendroctonus valens. Tras el ataque inicial del D. mexicanus, el pino infestado es invadido por el D. valens. Este insecto es muy conocido en los Estados Unidos de América, y sus costumbres se han recogido en boletines del Departamento de Agricultura de los Estados Unidos.4
[4 Hopkins, A. D., The Genus Dendroctonus, U.S.D.A. Bulletin N°. 83, 1909 : Beal, J. A., and Massey, C.L., Bark Beetles and Ambrosia Beetles, School of Forestry, Duke University, Durban N. Carolina, Bulletin 10, 1945; Keen, F. P., Insect Enemies of Western Forests U.S.D.A. Miscellaneous Publication N°. 273, 1938.]
Sin embargo, es interesante observar que, al parecer. hay un diámetro mínimo por debajo del cual no ataca al P. leiophylla de México Central. Numerosos datos registrados por el autor fijan este diámetro límite en 6 pulgs. (15 cms.), aproximadamente, medidos por fuera de la corteza al nivel del suelo. Tampoco aquí parece haber ningún período de letargo en el ciclo vital del insecto en esta zona, ya que se observó que los ataques se producían normalmente en todos los meses del año.
Ips bonanseai. Siguiendo a la invasión eficaz del árbol por el D. mexicanus y luego por el D. valens, viene otra de un tercer escólito, o sea, el Ips bonanseai Hopk. El insecto adulto descrito por Hopkins tiene un promedio de 3,3 mms. de longitud, es alargado, cilíndrico y de color pardo rojizo oscuro. Presenta una hendidura en el declive de los élitros y está armado de cuatro dientes cortos y agudos a cada lado. Esta especie no invade al árbol sino después de que los ataques del D. mexicanus han contenido por completo el flujo de resina, después de lo cual acometen solamente la parte superior del tronco y las ramas más grandes. El orificio de entrada atraviesa la corteza y el líber, hasta la madera. En el líber excava una cámara nupcial, de la que parten tres o cuatro galerías sinuosas de desove. Las hembras depositan los huevos en nichos espaciados alternativamente a los lados de estas galerías. Cada huevo, en su nicho, se recubre de excrementos, pero las galerías y la cámara nupcial quedan siempre abiertas. Al romper el huevo, la larva se abre camino alimentándose desde las galerías hasta el líber. Cuando la larva se ha desarrollado, construye una celda en el líber, donde pasa su período de crisálida. El nuevo adulto se abre camino a través del excremento que fué antes tejido de líber, hasta las galerías antiguas, y sale por los orificios de entrada y ventilación. No se intentó determinar el promedio del tiempo necesario para que este insecto complete su ciclo vital. Sin embargo, se observó que las infestaciones ocurrían durante los meses más fríos del año, lo que sugiere la ausencia del período de letargo en su ciclo vital.
Orden de ataque en las distintas zonas de la corteza del Pinus leiophylla.
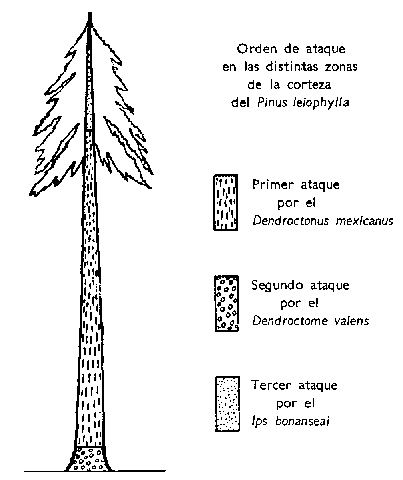
Importancia del Orden de Sucesión del Ataque en Arboles Independientes
La invasión del árbol vivo por estas tres especies de escólitos sigue un determinado orden de sucesión. El primer ataque corre invariablemente a cargo del D. mexicanus, y siempre, también, en el tronco del árbol si está lo bastante limpio, generalmente a una altura de 18 a 20 pies (5 ó 6 m.) del suelo. En los árboles más pequeños, o en los de muchas ramas bajas, el ataque suele empezar por el tronco, en la zona donde crecen las primeras ramas. A partir de este punto, la infestación se extiende hacia abajo, hasta una distancia de unas 18 pulgs. (46 cms.) del suelo y, al mismo tiempo, sube por el tronco, hasta un diámetro de 4 pulgs. (10 cms.), como máximo. Esta completa invasión del tronco por el D. mexicanus necesita, generalmente, de 7 a 10 días. En este momento, el D. valens invade con éxito el árbol desde el nivel del suelo hacia arriba, en una distancia aproximada de 18 pulgs. (46 cms.).
Tras la primera y segunda invasiones, el Ips bonanseai ataca la parte superior del tronco. En todos los casos se observó que el Ips ataca solamente cuando las hojas del pino empiezan a secarse, o sea, aproximadamente, a los 20 días de la invasión del tronco por el D. mexicanus. Ya entonces, el D. mexicanus ha alcanzado su estado de larva diminuta. La infestación del Ips baja por el tronco hasta un diámetro de 6 pulgs. (15 cms.), habiéndose observado también en la base de las ramas más grandes. Nunca se ha visto una inversión en este orden de ataque. Según muchas observaciones, parece ser que el ataque inicial por el D. mexicanus es absolutamente necesario para las invasiones sucesivas por el D. valens y el Ips bonanseai.
Ataques en Grupos
El comportamiento del D. mexicanus en grupo, al atacar los rodales de P. leiophylla es en todo semejante al de otras especies de Dendroctonus que atacan a las coníferas en el oeste de los Estados Unidos de América. Se ha seguido con bastante minuciosidad la propagación de varias infestaciones en pequeños rodales aislados, y en todos los casos la modalidad era igual. Primero fué atacado intensamente un pequeño grupo de árboles; luego los árboles vecinos fueron invadidos con mayor rapidez, hasta formarse un foco de infestación, que abarcaba de 10 a 20 árboles o más. Los árboles de los alrededores eran generalmente atacados por los nuevos escarabajos que emergían del primer grupo, propagándose la plaga con lentitud por todo el rodal en franjas cada vez más anchas. Una modalidad característica de estas pequeñas zonas atacadas es que en los bordes quedan aislados unos cuantos árboles supervivientes.
En otros casos, particularmente en los grandes rodales de pinos, los ataques en grupo se presentaban en mucho mayor escala. Varias de estas grandes infestaciones abarcaban de 5 a 10 o más acres (2-4 Ha.) y miles de pinos resultaban atacados simultáneamente. En cuanto surgían las crías de la primera generación, atacaban inmediatamente a los árboles vivos de las inmediaciones y, en un espacio de tiempo relativamente corto, un bosque de varios centenares de hectáreas quedaba completamente infestado. En algunos casos aparecían repentinamente grupos de gran extensión, en zonas no atacadas con anterioridad. Se cree que estas grandes infestaciones espontáneas son consecuencia de vuelos de insectos en masa desde una zona infestada donde el número de árboles vivos no basta ya como hospederos para la enorme multitud de nuevos escarabajos. El gran foco en grupo que se presentó en 1950 cerca de San Rafael se cree que se produjo de esta forma debido a que las nuevas generaciones de escarabajos del bosque infestado cerca de Amecameca no encontraron por los alrededores un número suficiente de árboles hospederos vivos. Este vuelo en masa cubría una distancia de 5 millas (8 Kms.), aproximadamente, pero en los rodales más pequeños, donde el ímpetu de la población no llega nunca a ser tan enorme, la acción de los escarabajos de nueva generación se limita por lo general a los pinos vivos más próximos. Mientras haya pinos vivos suficientes en los focos mayores de infestación, parece ser que los insectos se propagan, simplemente, partiendo del primer grupo atacante. La dispersión de los escarabajos por un pinar se sigue fácilmente por el cambio en la coloración de las hojas de los pinos infestados.
En el Cuadro 2 se indica el tiempo necesario para que se produzcan los cambios de coloración más importantes:
CUADRO 2
|
Coloración de las hojas |
No. de días aproximados desde la fecha del ataque |
Desarrollo medio del Dendroctonus mexicanus |
|
Descoloridas |
30 |
De larva pequeña a larva mediana, en líber |
|
Pardo |
60 |
Crisálida en la corteza exterior |
|
Rojo |
90 |
Aparición del adulto |
|
Negro (sin agujas) |
120 |
Aparición completa |
|
Cuerpos frutáceos del hongo Polyporus volvatus en el tronco |
150 |
|
|
Destrucción de la madera por el rápido desarrollo de los hongos. Arbol ya abatido |
300 |
|
Así, pues, la destrucción completa de un pino por el D. mexicanus se produce en menos de un año. El autor ha fotografiado sembrados de trigo entre tocones de pinos destruidos por el D. mexicanus hacía menos de un año.
Resistencia Comparativa del Pinus leiophylla y el P. montezumae var. macrocarpa.
En las zonas estudiadas se observó que el P. leiophylla era vencido rápidamente por los ataques del D. mexicanus y que en los centros epidémicos intensos eran pocos los árboles que sobrevivían.
El Profesor Martínez determinó las únicas otras especies de pinos en que se observó el ataque que fueron el Pinus montezumae y P. montezumae var. macrocarpa. En esta última especie, que a menudo se presenta, en rodales mixtos, con el P. leiophylla, al parecer, los escarabajos encontraban dificultades para atacar con éxito, y se descubrieron muchos árboles que habían sobrevivido a los violentos ataques. Se observaron muy pocos ataques contra el P. montezumae.
Las diferencias entre el P. leiophylla y el P. montezumae var. macrocarpa en cuanto a su resistencia al D. mexicanus, se manifestaron en el carácter y la abundancia de los tubos de resina, en la eficacia reproductora de los escarabajos y en la capacidad de la infestación epidémica de mantener su intensidad.
Diferencias en el Carácter de los Tubos de Resina. Los puntos de entrada de los escarabajos en el P. leiophylla se distinguían por el copioso flujo de oleorresinas. En general no se forman tubos de resina bien definidos sino que la resina tiende a gotear por fuera de la corteza, en largas regueras brillantes. En el P. montezumae var. macrocarpa los orificios de entrada son más numerosos y la resina se endurece rápidamente, formando tubos en forma de torre. Muchos de los adultos atacantes quedan atrapados por la rapidez con que se seca la resina y mueren en el tubo sin haber penetrado en el árbol. Esta diferencia de carácter entre los tubos de resina se debe a las que existen entre los componentes químicos de las oleorresinas que producen una y otra especie. El informe del Dr. N.C. Mirov revela que las oleorresinas del P. montezumae y del P. leiophylla son completamente distintas; la oleorresina del P. montezumae se compone casi exclusivamente de dextrosa alfa-pina, mientras que la resina del P. leiophylla contiene sólo 90 por ciento de dextrosa alfa-pina, 8 por ciento de dextrosa delta-3-careno y 1 por ciento de dextrosa sesqui-terpina.
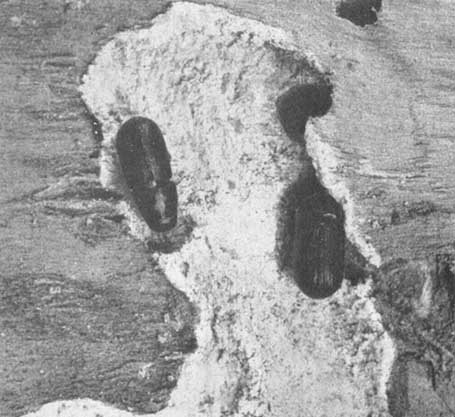
Las dos fotografías muestran el chorro de resina ocasionado por los ataques del Dendroctonus mexicanus:
- al P. leiophylla
Adultos de Ips bonanseai (x 3 aproximadamente) en galerías de desove parcialmente construidas,
Galería de desove y adulto (x 5) de Dendroctonus valens.
Diferencia en el Potencial Reproductivo del D. mexicanus. El D. mexicanus mata algunos árboles de P. montezumae var. macrocarpa, pero se ha observado, que en estos casos, era mayor el ataque por unidad de superficie de corteza, que en el P. leiophylla. Una comparación del número de puntos de entrada, de los cuales cada uno representa el ataque de un par de escarabajos padre, y de los puntos de salida, que representan la aparición de la progenie, nos demostrará esta diferencia.
El Cuadro 3 se basa en el recuento de los puntos de entrada y de salida en zonas de corteza elegidos al azar en dos árboles representativos de ambas especies de pino. Los dos árboles habían muerto por ataque del D. mexicanus.
CUADRO 3. - COMPARACIÓN DEI. NÚMERO DE PUNTOS DE ENTRADA Y DE SALIDA DEI, Dendroctonus mexicanus EN EL P. leiophylla Y EL P. montezumae var, macrocarpa
|
|
P. leiophylla |
P. montezumae var. macrocarpa |
||
|
Pies2 |
m.2 |
Pies2 |
m.2 |
|
|
Total de la superficie de corteza examinada |
6,8 |
0,6 |
5,5 |
0,5 |
|
Promedio del número de puntos de entrada por pie cuadrado (m.2) |
8,0 |
0,7 |
16,8 |
1,6 |
|
Promedio del número de puntos de salida por pie cuadrado (m.2) |
104,0 |
9.7 |
39,0 |
3,6 |
Este cuadro indica que el P. montezumae var. macrocarpa fué atacado por doble número de escarabajos que el P. leiophylla, en tanto que el número de crías en el P. leiophylla resultó dos veces y media mayor que en el P. montezumae var. macrocarpa. En suma, si admitimos dos adultos padres por cada punto de entrada, el número de escarabajos que atacan al P. montezumae var. macrocarpa por pie cuadrado es de 33,6 (362 por m.2), y de 39 (420 por m.2) el de los que luego salen, lo que sólo permite un aumento insignificante por generación. Si se tiene en cuenta la mortalidad de los nuevos escarabajos en el momento del vuelo y en el ataque contra árboles nuevos, cabe esperar una disminución en su población.
Efecto de las Especies Hospederas en las Poblaciones de D. mexicanus. Los resultados que figuran en el cuadro 3 están completamente de acuerdo con las observaciones sobre el terreno. En los rodales puros o casi puros de Pinus leiophylla de edad uniforme, el D. mexicanus se convierte en epidémico y se propaga con rapidez, hasta que todo el rodal queda prácticamente destruido. Las epidemias de D. mexicanus son menos violentas cuando el P. montezumae var. macrocarpas constituye una proporción apreciable del rodal, y donde se han encontrado elevados porcentajes de esta especie, no se ha observado ninguna epidemia.
LUCHA CONTRA EL D. MEXICANUS
En estos estudios no se intentó ningún experimento de lucha, porque no se disponía de tiempo sino para recoger datos biológicos y ecológicos. Sin embargo, disponemos de algunos datos tanto sobre la lucha natural como sobre la artificial que, por tanto, damos aquí.
El orden sucesivo de los ataques indica que el Ips y el D. valens no pueden mantener infestaciones epidémicas más que con un carácter secundario a la del D. mexicanus. Por eso, los esfuerzos en la lucha se pueden concentrar en el D. mexicanus para evitar la virulencia y propagación de epidemias como las que están en curso actualmente.
Lucha Biológica
Pájaros. En los pinares de los Estados Unidos de América es un hecho muy corriente que el pájaro carpintero se alimente de las crías del escarabajo Dendroctonus, que se encuentran en la corteza de los árboles infestados, y en ocasiones se considera un factor importante en la lucha natural contra este insecto. Los pájaros carpinteros que se alimentan del Dendroctonus brevicomis que ataca al pino Ponderosa llegan a destruir hasta el 75 por ciento de las crías. Sin embargo, no se ha apreciado ningún efecto del pájaro carpintero en las infestaciones del D. Mexicanus. Esto puede muy bien deberse a la gran escasez de estos pájaros, que en estas zonas parece que están a punto de exterminarse.
Por el contrario, el Dr. Bonansea señala que la reducción de las poblaciones de insectos parásitos y entomófagos por los pájaros constituiría uno de los motivos de las infestaciones epidémicas de escarabajos de la corteza. No existen pruebas de que los pájaros hayan desempeñado esa función en las recientes infestaciones.
Insectos Voraces. En los árboles plagados por el D. mexicanus y en torno a ellos se encontró cierta cantidad de escarabajos voraces. En su mayor parte, pertenecen a la misma especie que los que se alimentan del D. brevicomis y son, sin duda, importantes en la lucha biológica contra el D. mexicanus.
Enoclerus sphegens F. (F.), Thanasimus nigriventris, Themnochila virascens, todos determinados por E. A. Chapin.Cymatodera sp., cerca de Morosa (Lee.), determinado por O. L. Cartwright.
Se ha visto que los adultos de las especies arriba citadas se alimentan de los adultos del D. mexicanus y en la corteza se encontraron formas larvarias junto con las larvas del escarabajo. Aunque estos escarabajos voraces se presentan en cantidades, sus efectos en la lucha contra el escarabajo taladrador de corteza no han sido suficientes para dominar el rápido incremento de las recientes epidemias.
Insectos Parásitos. En las crías de D. mexicanus recogidas en las cortezas infestadas de P. leiophylla se encontraron adultos de calcídidos pequeños. Estos fueron determinados por el Dr. B.D. Burks del Museo Nacional de los Estados Unidos como Pachyceras sp. y Tomicobia sp. Por la naturaleza de las crías no se pudo establecer exactamente que estos insectos himenópteros sean parásitos del D. mexicanus, pero es posible que lo sean, dados los conocidos hábitos parasitarios de muchas especies que pertenecen a este grupo. Se criaron un pequeño número de los calcídidos adultos y su relativa escasez muestra que no constituyen un factor importante en el control biológico del escarabajo.
Métodos Directos de Lucha
Los métodos empleados en la lucha directa contra el D. brevicomis deben resultar igualmente eficaces contra el D. mexicanus. Estos métodos consisten en quemar la corteza infestada para destruir los insectos, o en rociarla con aceites tóxicos penetrantes hasta que quede bien saturada. En cualquiera de estos casos es conveniente apear el árbol antes de tratar la corteza, y cuando se quema esta última, se aconseja descortezar el tronco.
El ingeniero Humberto Ortega, Jefe del Departamento de Protección Forestal del Gobierno de México, informa que en 1950 se llevaron a cabo trabajos de lucha directa contra las plagas, en partes de la Unidad Forestal de San Rafael. En los terrenos de propiedad particular se consiguió que los propietarios proporcionaran la mano de obra para talar, podar y descortezar los árboles infestados. El ingeniero forestal de la Unidad marcaba los árboles que había que apear y dirigía la quema de las ramas y copas infestadas. A los propietarios se les permitió vender la madera para resarcirse de los gastos de esta operación. Varios grandes centros de infestación fueron tratados por el mismo procedimiento.
Al examinar estas mismas zonas un año después, no se observó ningún nuevo ataque en los alrededores de las ya tratadas. Estos resultados indican que los trabajos para extirpar las plagas del D. mexicanus, producen un efecto local bien definido, y que se pueden obtener buenos resultados si estos trabajos se llevan a cabo concienzudamente.
En los «ejidos»5 comprendidos en las tierras comunales de la Unidad Forestal, los árboles muertos se ceden a los propietarios comunales o «ejidatarios», de acuerdo con la Ley, y no se han podido recaudar fondos para llevar a cabo trabajos de lucha directa. Debido a ello, en las zonas forestales de las tierras egidales no se han controlado las plagas. La Unidad Forestal, sin embargo, mantiene un vivero forestal y está replantando algunas de las zonas devastadas.
[5 Tierras restituidas o concedidas a comunidades agrícolas en virtud de la reforma agraria emprendida en 1915.]
Lucha por Procedimientos Dasocráticos.
La regulación de cortas y la dasocracia pueden ofrecer la mejor solución para evitar las plagas de D. mexicanus en los rodales de pinos. En los rodales de P. leiophylla destruidos por epidemias recientes, el crecimiento de los árboles más viejos ha sido rápido en los primeros 35 a 40 años; pero, pasado este espacio de tiempo, se produjo un brusco retardo, como lo prueba la ausencia de los anillos anuales de los árboles. Durante este período de interrupción del crecimiento vegetativo fué cuando se presentó la epidemia del escarabajo taladrador. Este dato sugiere que en los rodales de P. leiophylla de edad uniforme que han pasado por el período de crecimiento rápido, una corta a mata rasa más o menos en el momento en que la vegetación empieza a detenerse eliminará parcialmente el riesgo de una epidemia de escarabajos, y permitirá utilizar por completo toda la madera antes de su deterioro. La replantación en estas zonas taladas de especies más resistentes, tales como el P. montezumae var. macrocarpa puede ser un método adecuado de control del escarabajo en aquellas zonas donde interese una producción continua de pinos
AGRADECIMIENTO
El autor desea expresar su gratitud a las numerosas personas que le han ayudado en la preparación de este estudio: al Sr. John M. Miller, Entomólogo del Laboratorio de Insectos Forestales de Berkeley, Oficina de Entomología y Cuarentena Vegetal de la Dirección de Investigaciones sobre Insectos Forestales, Secretaría de Agricultura de los Estados Unidos, por la aguda que le ha prestado sobre el terreno y por sus numerosas y útiles sugerencias y criticas durante la preparación del manuscrito al Dr. J. A. Beal, Jefe de la Oficina de Entomología y Cuarentena Vegetal de la Dirección de Investigaciones s sobre Insectos Forestales Secretaría de Agricultura de los Estados Unidos; a la Dra. Dorothy Parker, Especialista en Botánica al servicio del Programa Agrícola que está desarrollando la Fundación Rockefeller en México, quien ha examinado el manuscrito y ha hecho valiosas Sugerencias al Dr. N. T. Mirov, Químico de la Secretaría de Agricultura de los Estados Unidos de América, que presta sus servicios en la Estación Experimental de Bosques y Pastizales de California, Berkeley, California, por sus datos sobre análisis químicos de las resinas al Ingeniero Humberto Ortega, Jefe del Departamento Mexicano de Protección Forestal, por su información sobre las medidas que se aplican actualmente en México para combatir los insectos. Todas las fotografías fueron tomadas por el autor; el Sr. Byron MacLellan, Fotógrafo de la Fundación Rockefeller, hizo las reproducciones y ampliaciones necesarias.
Cupones de la UNESCO
La Organización de las Naciones Unidas para la Educación, la Ciencia y la Cultura (UNESCO) ha solicitado de UNASYLVA que ponga en conocimiento de sus lectores ciertos hechos relativos a los «Cupones de la UNESCO». El propósito de estos cupones fué, en un principio, facilitar a las instituciones y a los particulares de los países miembros de la UNESCO, la adquisición de libros suscripciones a periódicos y a otras publicaciones de distintos países, a pesar de la escasez de divisas extranjeras y de las restricciones en los cambios. Como consecuencia del éxito del plan de Cupones para Libros, se ha resuelto extender su empleo para facilitar, además, la adquisición de películas culturales, científicas y educativas, películas fijas, y película virgen, así como la adquisición de material científico, de laboratorio, etc.
Los Cupones de la UNESCO hacen posible la importación de estos artículos al margen de los reglamentos monetarios normales y, en muchos casos, con facilidades especiales de exención de derechos arancelarios.
Las personas que deseen datos más detallados, pueden conseguir los folletos «El Cupón UNESCO para Libros», «El Cupón UNESCO para Películas», y «El Cupón UNESCO para Materiales Científicos», así como la reciente «Circular Informativa sobre el Sistema de Cupones», escribiendo a la siguiente dirección:
Oficinas de Cupones de la UNESCO 19, Avenue Kleber, París 16, Francia.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}