![]()
![]()
![]()
M. MURRAY
Introduction
Criteria of trypanotolerance
Basis of mechanisms of trypanotolerance
Genetics of trypanotolerance
Factors affecting the stability of trypanotolerance
Susceptibility of trypanotolerant cattle to other diseases
Conclusion
References
In Africa, it has long been recognized that certain breeds of cattle are able to survive in tsetse-infested areas where other breeds rapidly succumb (Pierre, 1906; ILCA, 1979). Furthermore, in many instances under these circumstance they also appeared to be productive. This trait has been termed trypanotolerance and is generally attributed to the indigenous taurine breeds of cattle in West and Central Africa, namely the N'Dama and West African Shorthorn. However, it was not clear whether survival was due to the fact that such animals were less liable to become infected or to the fact that they were more "tolerant" to the effects of infection. Although it is not known whether trypanotolerant animals are less liable to tsetse attack, it has now been shown that where tsetse challenge is high, trypanotolerant breeds such as the N'Dama are as likely to become infected as trypanosusceptible breeds (Murray et al., 1981a). Thus, while it is of interest to determine what factors affect host attractiveness to tsetse, in the context of trypanotolerance attention must be focussed on the factors that permit animals to resist the effects of infection.
Major comparative investigations on the question of trypanotolerance have been carried out on cattle throughout Africa, e.g. in Nigeria (Desowitz, 1959; Roberts and Gray, 1973a, b), The Gambia (Murray et al., 1981a; reviewed by Murray et al., 1982), Senegal (Toure et al., 1978), Burkina Faso (reviewed by Roelants, 1986; Akol et al., 1986), Kenya (Njogu et al., 1985; Ismael et al., 1985; Paling et al., in press) and in the ILCA/ILRAD Trypanotolerance Network (1986a, b). The main breeds studied included Ayrshire, Friesian, Holstein, Hereford and their crosses, as well as indigenous African breeds such as Zebu, Boran, West African Shorthorn and N'Dama. Their ages have varied from one week to fifteen years and they were maintained in villages, ranches or experimental stations where major differences in nutrition must have occurred.
In these investigations, cattle were infected by syringe inoculation with bloodstream trypanosomes, exposed to field challenge, laboratory challenge with wild caught tsetse or with tsetse experimentally infected. Irrespective of the mode of infection, the outcome of each study consistently confirmed the superior resistance of the N'Dama (and the West African Short horn) and showed that the basis of this trait was associated with the capacity of these animals to develop less severe anaemia. Anaemia is a well recognised and inevitable consequence of trypanosome infections in domestic animals in general and cattle in particular (Hornby, 1921; Murray, 1974; Morrison et al., 1981). Maasai herdsman recognize that cattle in tsetse infested areas can "run out of blood." It has now been definitely established that the measurement of anaemia gives a reliable indication of the disease status (Murray, 1979) and productive performance (ILRAD, 1986; ILCA, 1986a, b) of trypanosome- infected cattle.
Furthermore, resistance of these breeds to anaemia appeared to be correlated with the ability to limit the intensity, pre- valence and duration of parasitaemia (Dargie et al., 1979). A similar capacity to control parasitaemia and resist anaemia has been shown by the Orma Boran cattle in Kenya (Ismael et al., 1985) and to an even greater extent by several species of wild Bovidae (Murray et al., 1981b).
A study which indicated that the height of parasitaemia per se can influence the degree of anaemia was a dose reponse experiment in which grouts of N'Dama and Zebu were inoculated sub cutaneously with 103, 105 or 108 bloodstream forms of Trypanosoma brucei (Murray et al., 1979). It was found that the number of parasites inoculated influenced not only the prepatent period but also the height of parasitaemia. Thus, the N'Dama and Zebu groups that received the largest doses had the shortest prepatent period and the highest parasitaemia and, correspondingly, developed the most severe anaemia. In confirmation of their trypanotolerant nature, the N'Dama were better able to control parasitaemia and developed less severe anaemia than the Zebu which received the same dose; thus, it was necessary to challenge the N'Dama with 5log10 more organisms to produce similar levels of parasitaemia and similar severity of anaemia as in Zebu. This work, using T. brucei in cattle, is the only dose response study of which we are aware. It has important implications both in relation to control of parasitaemia and development of anaemia. However, it is our impression with T. congolense infections in cattle that while infective dose determines the prepatent period, it does not affect the height of parasitaemia or the severity of anaemia. In the same way in mice, increasing doses of T. congolense (Morrison and Murray, 1985) or T. brucei (ILRAD, unpublished data) resulted in shorter prepatent periods, but there were no significant increases in the height of parasitaemia or the time to death after the first peak of parasitaemia.
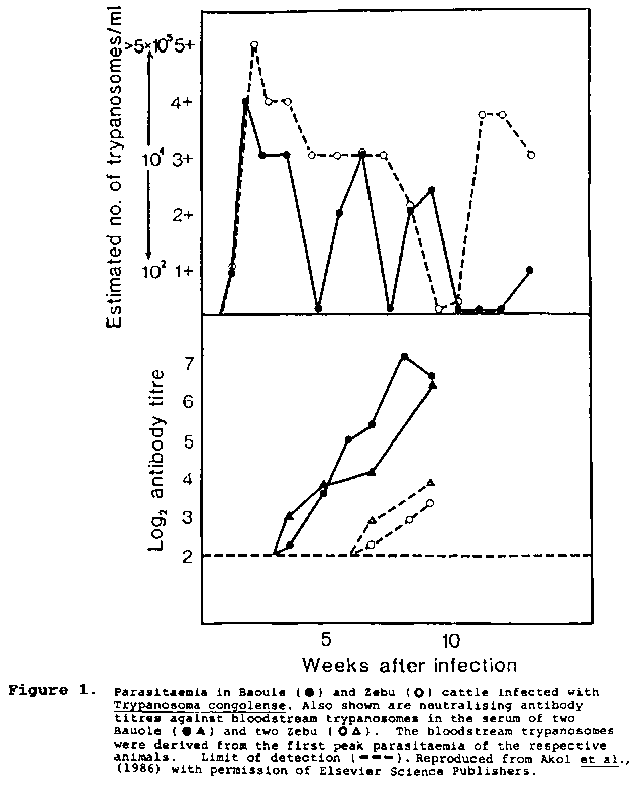
As parasitaemia wave remission is effected by antibodies directed against the surface coat antigens of the trypanosome (reviewed by Murray and Urquhart, 1977), it is generally assumed that the superior capacity of trypanotolerant animals to control parasitaemia is associated with a better immune response, al-though in cattle, at any rate, there are only a few preliminary studies in N'Dama infected with T. vivax (Desowitz, 1959) and Baoule (West African Shorthorn) infected with T. congolense (Akol et al., 1986) which suggested that this might be the case.
Thus, available evidence now indicates that resistance to trypanosomiasis is determined by at least three main characteristics, namely, the ability to control parasitaemia, the ability to develop an effective immune response and the ability to resist anaemia.
Regulation of parasitaemia and immune response
Considerable understanding of the relationship between susceptibility, parasite growth and the immune response has been provided by the work of Black et al. (reviewed in 1985) on the mouse model system. Mice vary in their susceptibility to T. brucei (Black et al., 1983), to T. congolense (Morrison et al., 1978) and to T. vivax (Mahan et al., 1986). Basically, it has been found that the more resistant strains of mice such as the C57Bl are better able to control parasitaemia and produce a superior antibody response. The differences in the immune response to infection could not be attributed to inherent differences in the immune response to the parasite, as no differences in antibody responses occurred between different strains of mice when immunized with non-dividing irradiated T. brucei (Black et al., 1983), T. congolense (Morrison and Murray, 1985) or T. vivax (Mahan et al., 1986). In T. brucei infections, it was concluded that the height of parasitaemia is controlled by the availability of molecules which maintain the parasites as dividing forms. When the molecules reach a limiting concentration, the parasites cease dividing and start to degenerate. In vitro analysis suggests that T. brucei growth promoting activity is associated with serum components of molecular mass greater than 100 kD (Black et al., 1985). The importance of parasite growth regulation in the control of parasitaemia is further emphasised by the findings which suggest that antibody responses against T. brucei are stimulated by fragments derived from senescent non-dividing organisms but not by actively dividing parasites (Sendashonga and Black, 1982). As with T. brucei, the elimination of T. vivax and T. congolense from the blood is antibody-mediated. However, there, would appear to be more important differences in the role played by antibody responses in controlling T. brucei, T. congolense and T. vivax parasitaemia. Antibody responses to T. brucei (Sendashonga and Black, 1982; Black et al., 1983) and T. congolense (Mitchell and Pearson, 1986; Whitelaw et al., 1983) parasitaemia occur after most organisms have differentiated to non-dividing forms, the stumpy form in the case of T. brucei and hence when parasite population expansion in the blood is no longer exponential. In contrast, antibody control of parasitaemia in C57B1/6 mice infected with T. vivax occurred during the exponential phase of parasite population increase in the blood (Mahan et al., 1986). Thus, the capacity to control T. brucei and T. congolense parasitaemia appears to lie in the ability to regulate parasite growth followed by the induction of the immune response, while in T. vivax infections it appears to be directly dependent on the ability to mount an immune response.
Another aspect of the inability of susceptible C3H/He mice to control parasitaemia following infection with T. brucei (Black et al., 1986) or with T. vivax (Mahan et al., 1986) resulted from an impaired capacity of parasite-induced antibody-containing cells to secrete immunoglobulin. Such cells regained the ability to secrete antibody within 24 hours after trypanosome elimination by treatment with the trypanocidal drug Berenil (Black et al., 1986; Mahan et al., 1986), suggesting that the block in antibody secretion was maintained by living parasites or short-lived components of degenerating parasites.
It was also possible to manipulate population growth rates in mice infected with T. congolense and T. brucei (Murray and Morrison, 1979) and with T. vivax (ILRAD, unpublished data). Mice treated with heat-killed Corynebacterium parvum prior to infection were able to control parasitaemia and survive for longer. The effect was most marked in strains of mice with the greatest innate resistance to trypanosome infection, e.g., the C57B1/6. Thus, sera from C. parvum treated mice contained a group of hydrophilic molecules (100 kD - 1000 kD) which competitively inhibited the capacity of foetal bovine serum (FBS) to support multiplication of T. brucei in vitro (ILRAD, unpublished data). Similar molecules were released by macrophages of C. parvum treated mice, by macrophages fed C. parvum in vitro and, at a 4000 fold lower efficiency, by C. parvum incubated in vitro in the absence of macrophages. Synthetic muramyldipeptide also competitively inhibited the capacity of FBS to support T. brucei multiplication in vitro, suggesting that the C. parvum inhibitory effect is mediated by fragments of peptidoglycan. The competitive interaction between the growth-promoting factors in FBS and the growth-inhibiting factors produced by C. parvum might indicate a competitive interaction for a receptor site at the level of the trypanosome, a receptor usually reserved for the binding and uptake of serum nutrients which regulate multiplication.
The identification and characterization of the factor (s) that promote and inhibit parasite growth, as well as the trypanosome-factors that inhibit immunoglobulin secretion in trypanosusceptible animals, could lead to new therapeutic or immunological approaches to prevent or control parasite growth. Understanding prevention could result in a vaccine; better control might make it possible to turn the trypanosome status of a Zebu into that of an N'Dama and of an N'Dama into that of a wild Bovid.
As discussed, only limited data are available on the immune responses of trypanotolerant and trypanosusceptible cattle to trypanosome infections. Superior antibody responses to T. vivax infections have been reported in N'Dama (Desowitz, 1959) and in Baoule cattle infected with T. congolense (Akol et al., 1986). Desowitz (1959) suggested that the basis of this superiority was a better innate capacity to mount a secondary immune response. While no differences were found in antibody responses between trypanotolerant and trypanosusceptible cattle to infecting T. congolense metacyclic parasites delivered by tsetse (Baoule v Zebu, Akol et al., 1986; N'Dama v Boran, Paling, ILRAD personal communication) or to syringe inoculated forms of T. brucei (Baoule v Zebu, Pinder et al., 1984), Akol et al. (1986) showed that the Baoule mounted a much greater neutralising antibody response to the first peak bloodstream trypanosomes (Figure 1), possibly reflecting an inherent ability to produce a superior secondary response. In the same way, the higher number of B lymphocytes found in N'Dama compared with Boran, prior to and throughout the course of infection with T. congolense (Ellis et al., 1987), might be due to the fact that the animals used in that study had experienced a previous infection with an unrelated serodeme of T. congolense, possibly giving the N'Dama the opportunity to prime their superior immune system. In this respect, Saror et al. (1981) found no difference in total blood leukocytes in N'Dama and Zebu cattle with no previous exposure to trypanosomiasis.
Figure 1. Weeks after infection
There is evidence to indicate that the macrophage could play an important role in controlling parasitaemia Thus, it was found that wildebeest (Connochaetes taurinus) infected with T. brucei were better able to control parasitaemia than their cattle counterparts (Zebu x Charolais) and, unlike cattle, did not develop anaemia (Rurangirwa et al., 1986). Both wildebeest and cattle produced similar IgM, IgG1 and IgG2 antibody responses to the infecting trypanosome. However, it was found that serum from infected wildebeest had a higher capacity than cattle to induce adherence of trypanosomes to their own peripheral blood leukocytes (PBL) and that this difference could be attributed to the presence of IgM receptor (s) in wildebeest PBL, which were not present in the PBL of the cattle studied. Such a mechanism could account for the lower parasitaemia encountered in trypanotolerant animals. The cattle in this study were trypano-susceptible Zebu x Charolais and it is important that trypanotolerant N'Dama cattle be investigated in the same way.
Haemopoietic responsiveness
There is little doubt that the intensity of parasitaemia has a significant effect on the severity of the subsequent anaemia, an observation that has led to the general conclusion that the superior resistance of trypanotolerant animals lies in their ability to control parasitaemia rather than in an inherent capacity to resist red cell destruction or mount a more efficient erythropoietic response (Dargie et al., 1979).
However, differences in the severity of anaemia have been observed under circumstances where no difference in the intensity or prevalence of parasitaemia occurs.
Thus, a number of workers have confirmed that calves of less than one year old are more resistant than adults to the effects of trypanosomiasis (Fiennes, 1970; Maxie and Valli, 1979; Wellde et al., 1981; Murray et al., 1982). While Wellde et al. (1981) concluded that parasitaemia was possibly less severe in calves infected with T. congolense, in an experiment carried out at ILRAD on Boran cattle aged six weeks and eighteen months, the calves developed anaemia that was significantly less severe than in adults, despite the fact that no significant differences could be quantified either in the intensity or prevalence of the parasitaemia. These results might indicate that younger animals have a superior erythropoietic response. In this respect, it is of interest that calves are more resistant than adults to babesiosis (Callow et al., 1976; Levy et al., 1982).
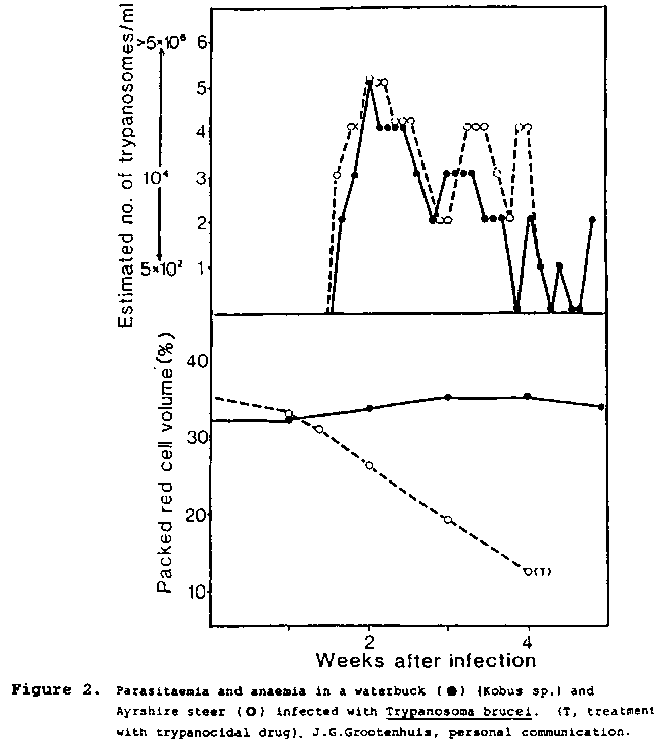
Certain species of wild Bovidae can develop high levels of parasitaemia following trypanosome infection, e.g., waterbuck (Kobus sp.) infected with T. brucei. Despite this, no significant changes in red cell numbers occurred, in contrast to the severe anaemia in a trypanosusceptible cow with equivalent levels of parasitaemia (Figure 2). These findings might suggest that the waterbuck red cells were more resistant to the destructive effects of the trypanosome.
In probably the most informative experimental study to date on the question of trypanotolerance, Paling et al. (in press) presented results on N'Dama and Boran cattle which indicated that the superior capacity of the N'Dama to control parasitaemia and resist anaemia are processes that are controlled genetically but are not directly linked to each other. The eight N'Dama used in the study were of Gambian origin and had been obtained by intrauterine implantation of embryos into Boran female cattle at ILRAD, Nairobi. The N'Dama along with corresponding Boran controls had been challenged on four occasions with Glossina morsitans centralis infected with clones of T. congolense belonging to four different but equally virulent serodemes. To prevent death, any infected animal with a PCV value of 15% or less was treated with Berenil. In addition, all infections were terminated with Berenil on 164, 127, 149 and 133 days after each challenge.
Figure 2. Weeks after infection
No N'Dama required treatment, but 77% of all infections in the Boran (32 animals) did. Moreover, in contrast to the severe weight losses experienced by trypanosome-infected Boran, compared with uninfected controls, trypanosome infection did not appear to affect body weight or liveweight gains in the N'Dama. Also in contrast to the Boran, infected female N'Dama continued to show normal oestrus-cycle activity.
There were no significant differences between either the breed groups or among the infections of the four T. congolense clones in the prepatent period or in the number of days to the first peak in the parasitaemia However, the average number of parasites per ml blood at the first parasitaemia peak was slightly but significantly lower in N'Dama than in Boran. The overall mean parasitaemia score of the N'Dama during the four infection periods was significantly lower than the overall mean score of untreated Boran. Within the N'Dama group, individual animals demonstrated significantly higher or lower mean parasitaemia levels and this trend was consistent throughout all four infections.
The kinetics of the anaemia in the group of N'Dama cattle during each of the four infection periods was very similar. First, there was a short phase during the development of parasitaemia from day 10 to 15 when the average daily PCV value in N'Dama and Boran dropped at the same rate. This was followed by a period from day 15 to 35, during which the average PCV in N'Dama decreased, but at a slower rate than that of the Boran. Might this suggest that N'Dama red cells are more resistant to destruction? From day 35-60, the average daily PCV value in N'Dama remained at the same level but most of the Boran required treatment. The lowest average PCV value reached by the N'Dama varied between 22.5 and 23% during the first two infections and between 25 and 27% during the second two infections. Next was a period of spontaneous recovery in the N'Dama, as assessed by a return to an average PCV value of 30%. In contrast, no spontaneous recovery occurred in the Boran. Most of the Boran required treatment and those that did not maintained a persistent average PCV value of 22%. The overall mean PCV value of the N'Dama was 29.2% during the four infection periods and this was significantly higher than the mean of 24.7% for the untreated Boran.
While the mean PCV value of the N'Dama group increased from 27.9% during the first infection to 31.2% during the fourth infection, there was no corresponding decrease in trend in the parasite numbers measured during the four infections. In the same way, some N'Dama demonstrated consistently higher PCV values during all four infections while others showed better parasite control. Thus, there was no correlation in the N'Dama between the level of parasitaemia, expressed as the mean parasitaemia score during the period of infection and the level of anaemia expressed as the mean PCV.
The superior resistance of N'Dama compared with Zebu to trypanosome infection was once again confirmed by this study and the same general conclusion made, namely, that N'Dama exhibited a superior capacity to control parasitaemia and resist anaemia. However, when mean parasitaemia and PCV values of the N'Dama were computed over four infection periods, no direct correlation could be established either at a group or individual level, indicating that both processes although controlled genetically are not directly linked to each other.
While control of parasite growth would still appear to be an important component of trypanotolerance, other explanations must be sought for the capacity of certain groups of cattle and individuals within each group to resist anaemia. One interesting possibility has been reported by Esievo et al. (1986) who measured erythrocyte surface sialic acid concentrations in uninfected N'Dama and Zebu with no previous exposure to trypanosome. The sialic acid concentrations in the N'Dama were about seven-fold greater than in the Zebu, although no differences in red cell surfaces of the N'Dama are stronger than the Zebu and that, if the presence in trypanosomes of the sialic acid-cleaving enzymes, sialidase (Esievo, 1983) or proteases (Londsdale-Eccles and Grab, 1986) is significant, sialic acid concentrations on red cell surface might play an important role in host resistance to trypanosome-induced anaemia.
In addition, in the study of Paling et al. (in press), there was evidence that the N'Dama had a greater inherent capacity to make better tertiary erythropoietic responses. Thus, the lowest average daily PCV values reached by the N'Dama group were 22.5, 23, 25 and 27% in consecutive infections, while the overall mean of all PCV values covering each of the four infection periods was 27.9, 28.7, 29.1 and 31.2%. Both results show a significant upward progression. This improved responsiveness was not observed in the Boran subjected to four consecutive infections and was not related to improvement in control of parasitaemia or to a difference in virulence among the four clones of T. congolense used.
Physiological adaptation
Another aspect of trypanotolerance which could be important and must be considered is that trypanotolerant cattle are more resistant to constraints in production because of superior physiological adaptation to factors such as food utilization, heat tolerance and water conservation. Unfortunately, critical observations concerning these qualities are not available at present. However, it is known from work on other breeds of cattle that genetically determined differences in voluntary food intake do exist. These have been ascribed to variations in fasting metabolism and maintenance energy requirements (Vercoe, 1974; Frisch and Vercoe, 1978). Where food intake is marginal, as it is in many areas of Africa, breeds with an inherently high maintenance requirement will suffer most, as demonstrated by weight loss or reduced weight gains. Thus, it is possible that trypanotolerant breeds such as N'Dama, especially when infected, are better adapted to remain productive in the prevailing conditions in Africa.
Little is known about water conservation and heat tolerance in trypanotolerant breeds. Pagot (1974) has pointed out that N'Dama can withstand higher levels of humidity than Zebu. In addition, there is a considerable variation in the rectal temperature of N'Dama during the course of the day in The Gambia, with temperatures ranging from 34.4°C at dawn to 41.1°C in late afternoon (Greig and McIntyre, 1979). The teleological argument for such a phenomenon is that thermoregulation under cold conditions, i.e., below 20°C, for tropical breeds of cattle requires the use of body energy stores. Thus, when caloric intake is low, energy is conserved by allowing the body temperature to fall (D. Robertshaw, personal communication). With regard to thermoregulation more is known about the zebu type breeds. Zebu are more heat tolerant than imported Bos taurus, either because their simple sac-like sweat glands and shallow hair follicle depth help promote sweating (Jenkinson and Nay, 1973) or because their lower maintenance requirements produce less heat load. Amakiri (1974) examined the histological skin structure of Nigerian breeds of cattle including N'Dama, Muturu and White Fulani and found that, on the basis of sweat gland size, the N'Dama were more akin to the exotic Bos taurus and, by inference, were less well-adapted to the tropical Nigerian environment. However, until sweating rate is actually measured, this conclusion is only speculative (D. Robertshaw, personal communication). The other characteristics of the skin, namely follicle depth and hair length, suggested that all three of these indigenous African breeds had the skin type associated with successful habitation of warm climates (Amakiri, 1974).
It has also been considered that the skin might play a role in trypanotolerance by affecting transmission. Thus, in wild Bovidae, chancres, the localised skin reactions that develop in cattle following the successful feed of trypanosome-infected tsetse, appeared less frequently and were smaller than in susceptible cattle subsequent to which the prepatent period was longer and the height of parasitaemia was lower (Murray et al., 1981b). Also, while Akol et al. (1986) reported possible differences in chancre reaction in Zebu and trypanotolerant taurine cattle in West Africa, no difference in prepatent periods occurred. Furthermore, no differences in chancre development or in prepatent period occurred between N'Dama and Boran following infection with T. congolense transmitted by Glossina morsitans centralis (ILRAD, 1985), demonstrating that the chancre cannot be used as an indicator of trypanotolerance in cattle.
Studies in East Africa, where the indigenous breed is the East African Zebu, have shown that its water requirement is about half that of the Hereford and is similar to that of several species of wild Bovidae, such as the eland (EAVRO, 1967). Zebu were also better able to conserve evaporative and faecal water than Hereford. Zebu deprived of water stopped eating and metabolized fat, with a resultant reduction of urinary and faecal water losses. Zebu could form faeces as dry as 110g water to 100g dry matter, whereas Hereford were unable to form faeces containing less than 300g water to 100g dry matter. This capacity for conserving water was inherited as a dominant trait in Zebu-Hereford crosses. It is likely that N'Dama have adapted to an even greater extent than Zebu because trypanosome-free N'Dama turn over less water than Zebu in terms of ml per kg bodyweight (Dargie, 1980).
It has now been definitively established that the capacity of certain breeds of cattle such as the N'Dama to resist the effects of trypanosome infection and maintain essential physiological functions is an innate trait, although, as will be discussed later, it can be affected by environmental influences. However, no estimates of heritability of trypanotolerance are available (Dolan, 1987). What results do exist suggest that trypanotolerance is inherited as a dominant trait. Stewart (1951) reported crossbreeding studies involving N'Dama, Zebu and Ghanaian Shorthorn (a trypanotolerant genetic mix) in which he produced a larger, more productive animal that retained its resistance to trypanosomiasis. Similarly, Chandler (1952) found that N'Dama/Zebu crossbreds retained a significant degree of trypanotolerance when exposed to natural tsetse challenge. Nine of twelve Zebu exposed died of trypanosomiasis, but only four of twelve crosses died; all eight N'Dama were alive and in good condition at the end of the experiment. In breeding experiments in Cote d'Ivoire, involving large numbers of N'Dama and Jersey, Letenneur (1978) found that the F1 cross produced an excellent animal as regards growth and milk production. It was stated that such crosses retained their tolerance, although no information was given on the level of fly challenge or on the prevalence of trypanosomes. However, crossbreds with greater than 50% Jersey background appeared to be less hardy and gave equivocal results.
A series of studies have been carried out on a range of inbred strains of mice, exhibiting a spectrum of susceptibility to T. congolense (Morrison and Murray, 1979; Pinder, 1984) and to T. rhodesiense (Levine and Mansfield, 1981; Greenblatt et al., 1984). The underlying genetic basis of susceptibility was examined in F1 hybrids and backcrosses derived from mouse strains of high and low susceptibility, while the influence of H-2 haplotype was evaluated using H-2 congenic strains of mice. The general conclusions of these studies were that resistance was inherited as a dominant trait, that it represented a polygenic model of inheritance and that it was not under the influence of H-2 haplotype. However, it should be emphasized that the mouse infections with high parasitaemias and inevitable death may not be a good model-system for cattle. In the same way, while no critical studies have been done to compare the degree of anaemia that develops in strains of mice of different susceptibility, it is my impression that, unlike cattle, the differences are minor.
A major constraint to establishing estimates of heritability of trypanotolerance in cattle has been related to the difficulty of defining and measuring the character, i.e., what precisely is trypanotolerance? However, in this respect there is convincing evidence that the severity of anaemia, as measured by PCV, in cattle in tsetse-infested areas is strongly correlated with production traits, such as reproductive performance and growth (ILCA, 1986a, b) and this has now been confirmed under controlled experimental conditions at ILRAD (ILRAD, 1986). Thus, PCV values induced during the course of a trypanosome infection might serve as a selection criterion for trypanotolerance. Nevertheless, the availability of a genetic marker for trypanotolerance would be ideal, as it would permit assessment without recourse to infection. However, to date such a marker has not been identified. Probably the best candidate at present is the major histocompatibility complex (MHC). This functions to direct and control immune responses and it is known that individuals with certain MHC are more likely to suffer from certain diseases. Thus, an investigation of the association between trypanotolerance and particular MHC gene products might prove profitable.
Several stress factors have been incriminated as affecting the susceptibility of trypanotolerant cattle to the effects of trypanosomiasis. Thus, it was believed by some that the resistance of trypanotolerant breeds was largely the result of acquired immunity to local trypanosome populations and that "tolerance" would break down if cattle were moved. There is evidence to confirm that trypanotolerant cattle, exposed to tsetse challenge, do become more resistant, either as a result of locally acquired immunity (Murray et al., 1982) or a primed erythropoietic response (Paling et al., in press). While exposure to new strains of trypanosomes will undoubtedly lead to infection, the superior genetic resistance of trypanotolerant breeds will ensure that their chances of survival and acquiring resistance in new locations will be significantly greater than for trypanosusceptible breeds. This conclusion has been confirmed by several reports which describe the movement of trypanotolerant cattle over large distances (Stewart, 1951; Ferguson, 1967) and their success-fur introduction into tsetse areas where other breeds cannot survive. For example, the Dahomey (Lagune) breed was first -established in Zaire from West Africa in 1904 and the N'Dama in 1920 (Mortelmans and Kageruka, 1976) and much more recently N'Dama were introduced into the Central African Republic, Gabon and Congo (ILCA, 1979).
Other stresses believed to be involved include, overwork, intercurrent disease, repeated bleeding, as well as pregnancy, parturition, suckling and lactation. Probably one of the most important factors is nutritional status of the host. Thus, in arid conditions when fodder is in short supply, N'Dama cattle have to trek many miles in order to obtain sufficient food. Under these circumstances, infected cattle even with only moderate anaemia are less able to cope and their capacity to forage and obtain sufficient food is compromised. However, it has been found that when such village N'Dama in The Gambia were supplemented even with only small amounts of groundnut meal, a local by-product, not only was their growth rate improved but they were significantly less anaemic than non-supplemented animals and did not forage so widely (W.I.M. McIntyre, personal communication). These exciting observations need urgent investigation.
A major question to be addressed is to what extent the degree of tsetse-trypanosomiasis risk affects the performance of trypanotolerance cattle. Preliminary field observations indicate that as the risk increases, production falls (ILCA, 1979). However, it is not known to what extent environmental/management factors might have influenced these findings and critical studies are required to address this question.
Another possibly important aspect of trypanotolerance is that it has been shown, at least in mice, that certain trypanocidal drugs, e.g., difluoromethylornithine, are more efficacious in animals which are innately more resistant and whose immune system is intact (De Gee et al., 1983; Bitonti et al., 1986). Therefore, if trypanocidal drugs are being considered for use in trypanotolerant cattle, it is interesting to speculate that not only would the number of treatments required be less but each treatment might be more effective.
While in most instances detailed critical evidence is lacking, our discussions with workers in the field and our own experience has convinced us that the taurine trypanotolerant breeds of West Africa are resistant not only to trypanosomiasis but also to several other important infectious diseases. Of major significance are the reports that both the N'Dama and West African Shorthorn are resistant to streptothricosis (Stewart, 1937; Coleman, 1967; Oduye and Okunaiga, 1971). Furthermore, N'Dama appear to be more resistant to tick-borne diseases, including heartwater (Cowdria ruminantium), anaplasmosis and babesiosis (Epstein, 1971). These observations might indicate a greater resistance to ticks per se. N'Dama would also seem to possess some degree of resistance to helminthiasis (A.A. Ilemobade, personal communication). In this respect, the Red Maasai sheep of Kenya have been shown to be significantly more resistant than other breeds not only to trypanosomiasis (Griffin and Allonby, 1979a, b) but also to haemonchosis (Preston and Allonby, 1979). On the other hand, N'Dama and West African Shorthorn are reputed to be much more susceptible to rinderpest than Zebu (Stewart, 1937, 1951; Cornell and Evans, 1937; Ferguson, 1967). Van Hoeve (1972) noted that Muturu, but not N'Dama, were highly susceptible to footrot.
While the foregoing observations require confirmation and more detailed investigation, it would appear that the N'Dama and West African Shorthorn possess a unique genetic advantage that makes them more resistant not only to trypanosomiasis but also to some of the other major infectious diseases of Africa.
Through a process of rigorous natural selection, Africa has provided her own answer to the problem of disease control in domestic livestock. Breeds of cattle are now known to exist that possess a significant degree of resistance to tsetse-transmitted trypanosomiasis, as well as to several other important infectious diseases. Moreover, it has been shown that these animals are productive and, with proper management, have considerable potential. Recognition that the degree of anaemia induced during a primary trypanosome infection reflects the trypanotolerant status of the host could allow estimates of heritability to be made and rational breeding programmes to be instituted.
Major research priorities must be to identify the mechanisms that regulate parasite growth, allow the development of an effective immune response and prevent pathogenic degrees of anaemia. An understanding of the mechanisms responsible might permit their manipulation and lead to novel strategies for the control of trypanosomiasis by therapeutic or immunological means, or even by molecular genetics. At the same time, marker (s) might be identified for genetic resistance, permitting the selection of breeding stock without having to infect animals.
Such research must be strongly encouraged in view of the poor prospects for the development of a vaccine or new trypanocidal drugs.
Akol, G.W.O., E. Audie, M. Pinder, S.K. Moloo, G.E. Roelants and Max Murray. 1986. Susceptibility and immune responses of Zebu and taurine cattle of West Africa to infection with Trypanosoma congolense transmitted by Glossina morsitans centralist Vet. Immunol. Immunopathol. 11: 361-373.
Amakiri, S.F. 1974. Sweat gland measurements in some tropical and temperate breeds of cattle in Nigeria. Anim. Prod. 18: 285-291.
Bitonti, A.J., P.P. McCann and A. Sjoerdsma. 1986. Necessity of antibody response in the treatment of African trypanosomiasis with -difluoromethylornithine. Biochem. Pharmacol. 30: 331-334.
Black, S.J., C.N. Sendashonga, P.A. Lalor, D.D. Whitelaw, R.M. Jack, W.I. Morrison and Max Murray. 1983. Regulation of the growth and differentiation of Trypanosoma (Trypanozoon) brucei brucei in resistant (C57B1/6) and susceptible (C3H/He) mice. Parasite Immunol. 5: 465-478.
Black, S.J., C.N. Sendashonga, C.O. O'Brien, N.R. Borowy, M. Naessens, P. Webster and Max Murray. 1985. Regulation of parasitaemia in mice infected with Trypanosoma brucei. In: Current Topics in Microbiology and Immunology. Berlin-Heidelberg: Springer-Verlag. 117: 93-118.
Black, S.J., C.N. Sendashonga, P. Webster and S.Z. Shapiro. 1986. Regulation of parasite-specific antibody responses in resistant (C57B1/6) and susceptible (C3H/He) mice infected with Trypanosoma (trypanozoon) brucei brucei. Parasite Immunol. 8: 334-352.
Callow, L.L., F.R. Emerson, R.J. Parker and S.G. Knott. 1976. Infection rates and outbreaks of disease due to Babesia argentina in unvaccinated cattle on 5 beef properties in south-eastern Queensland. Austral. Vet. J. 52: 446-450.
Chandler, R.L. 1952. Comparative tolerance of West African N'Dama cattle to trypanosomiasis. Ann. Trop. Med. Parasitol. 46: 127-134.
Coleman, C.H. 1967. Cutaneous streptothricosi of cattle in West Africa. Vet. Rec. 81: 251-254.
Cornell, R.L. and S.A. Evans. 1937. On the value and limitations of tissue vaccines against rinderpest. J. Comp. Path. 50: 122-135.
Dargie, J.D., P.K. Murray, Max Murray, W.R.I. Grimshaw and W.I.M. McIntyre. 1979. Bovine trypanosomiasis: the red cell kinetics of N'Dama and Zebu cattle infected with Trypanosoma congolense. Parasitol. 78: 271-286.
Dargie, J.D. 1980. Pathophysiology of trypanosomiasis in the bovine. In: Isotope and Radiation Research on Animal Diseases and Their Vectors. International Atomic Energy Agency, Vienna. IAEA-SM-240/28. pp. 121-131.
De Gee, A.L.W., P.P. McCann and J.M. Mansfield. 1983. Role of antibody in the elimination of trypanosomes after DL - difluoromethylornithine chemotherapy. J. Parasitol. 69: 818-822.
Desowitz, R.S. 1959. Studies on immunity and host parasite relationships. I. The immunological response of resistant and susceptible breeds of cattle to trypanosomal challenge. Ann. Trop. Med. Parasitol. 53: 293-313.
Dolan, R.B. 1987. Genetics and trypanotolerance. Parasitol. Today. 3: 137-143.
EAVRO. 1967. Annual Report: Physiology Section, Animal Production Division, East African Veterinary Research Organization. East African Community Services Organization, Nairobi. The Government Printer, Entebbe, Uganda. pp. 45-65.
Ellis, J.A., J.R. Scott, N.D. MacHugh, G. Gettinby and W.C. Davis. 1987. Peripheral blood leucocyte subpopulation dynamics during Trypanosoma congolense infection in Boran and N'Dama cattle: an analysis using monoclonal antibodies and flow cytometry. Parasite Immunol. 9: 363-378.
Epstein, H. 1971. The Origin of the Domestic Animals of Africa. Vols. 1 and 2. New York: Africana.
Esievo, K.A.N. 1981. In vitro production of neuraminidase (sialidase) by Trypanosoma vivax. International Scientific Council for Trypanosomiasis Research and Control. 16th Meeting. Yaounde, Cameroon, 1979. OAU/STRC. Publication No. 111. pp. 205-210.
Esievo, K.A.N., D.I. Saror, M.N. Kolo and L.O. Eduvie. 1986. Erythrocyte surface sialic acid in N'Dama and Zebu cattle. J. Comp. Path. 96: 95-99.
Ferguson, W. 1967. Muturu cattle of Western Nigeria: II. Survivability, reproductive and growth performances in an areas of light Glossina palpalis density. J. West African Sci. Ass. 12: 37-44.
Fiennes, R.N.T. -W. 1970. Pathogenesis and pathology of animal trypanosomiases. In: The African Trypanosomiases. H.W. Mulligan, ed. George Allen and Unwin/Ministry of Overseas Development, London. pp. 729-750.
Frisch, J.E. and J.E. Vercoe. 1978. Utilising breed differences in growth of cattle in the tropics. World Anim. Rev. 25: 8-12.
Greenblatt, H.C., C.L. Diggs and D.L. Rosenstreich. 1984. Trypanosoma rhodesiense: analysis of the genetic control of resistance among mice. Infect. Immun. 44: 107-111.
Greig, W.A. and W.I.M. McIntyre. 1979. Diurnal variation in rectal temperature of N'Dama cattle in The Gambia. Brit Vet. J. 135: 113-118.
Griffin, L. and E.W. Allonby. 1979a. Trypanotolerance in breeds of sheep and goats with an experimental infection Trypanosoma congolense. Vet. Parasitol. 5: 97-105.
Griffin, L. and E.W. Allonby. 1979b. Studies on the epidemiology of trypanosomiasis in sheep and goats in Kenya. Trop. Anim. Hlth. Product. 11: 133-142.
Hornby, H. E. 1921. Trypanosomes and trypanosomiasis of cattle. J. Comp: Path. 34: 211-240.
ILCA. 1979. Trypanotolerant Livestock in West and Central Africa. Monograph No. 2. ILCA, Addis Ababa, Ethiopia.
ILCA/ILRAD. 1986a. The ILCA/ILRAD Trypanotolerant Network. Situation report, December 1985. Proceedings of a Network Meeting held at ILCA, Nairobi. June 1986. Addis Ababa, Ethiopia.
ILCA/ILRAD. 1986b. The African Trypanotolerant Livestock Network. Indications from Results. 1983-1985. December, 1986. ILCA Addis Ababa, Ethiopia.
ILRAD. 1985. Annual Report of the International Laboratory for Research on Animal Diseases, P.O. Box 30709, Nairobi, Kenya.
ILRAD. 1986. Annual Report of the International Laboratory for Research on Animal Diseases, P.O. Box 30709, Nairobi, Kenya.
Ismael, A.A., A.R. Njogu, G. Gettinby and Max Murray. 1985. Susceptibility of Orma and Galana Boran Cattle to infection with bloodstream forms of Trypanosoma congolense and T. vivax. International Scientific Council for Trypanosomiasis Research and Control. 18th Meeting. Harare, Zimbabwe, 1985. OAU/STRC. Publication No. 113. pp. 176-181.
Jenkinson, D. McE. and T. Nay. 1973. The sweat glands and hair follicles of Asian, African and South American cattle. Austral. J. Biol. Sci. 26: 259-275.
Letteneur, L. 1978. Crossbreeding N'Dama and Jersey cattle in Ivory Coast. World Anim. Rev. 27: 36-42.
Levine, R.F. and J.M. Mansfield. 1981. Genetics of resistance to African trypanosomes: role of the H-Z locus in determining resistance to infection with Trypanosoma rhodesiense. Infect. Immun. 34: 513-518.
Levy, M.G., G. Clabaugh and M. Ristic. 1982. Age resistance in bovine babesiosis: role of blood factors in resistance in Babesia bovis. Infect. and Immun. 37: 1127-1131.
Londsdale-Eccles, J.D. and Grab, D.J. 1986. Proteases in African trypanosomes. In: Cysteine Proteinases and Their Inhibitors. V. Turk ed. Berlin: Walter de Gruyter. pp. 189-197.
Mahan, S.M., L. Hendershot and S.J. Black. 1986. Control of trypanodestructive antibody responses and parasitaemia in mice infected with Trypanosoma (Duttonella) vivax. Infect. Immun. 54: 213-221.
Maxie, M.G. and V.E.O. Valli. 1979. Pancytopenia in bovine trypanosomiasis. In: Pathogenicity of Trypanosomes. G. Losos and A. Chouinard, eds. IDRA No. 132e. pp. 135-136.
Mitchell, L.A. and T.W. Pearson. 1986. Antibody responses in resistant and susceptible inbred mice infected with Trypanosoma congolense. Immunol. 57: 297-303.
Morrison, W.I., G.E. Roelants, K.S. Mayor-Withey and Max Murray. 1978. Susceptibility of inbred strains of mice to Trypanosoma congolense: correlation with changes in spleen lymphocyte populations. Clin. and Exp. Immunol. 32: 25-40.
Morrison, W.I. and Max Murray. 1979. Trypanosoma congolense: inheritance of susceptibility to infection in inbred strains of mice. Exp. Parasitol. 48: 364-374.
Morrison, W.I., Max Murray and W.I.M. McIntyre. 1981. Bovine trypanosomiasis. In: Diseases of Cattle in the Tropics. Economic and Zoonotic Relevance. M. Ristic and I. McIntyre eds. Current Topics in Veterinary Medicine and Animal Science. Vol. 6. The Hague: Martinus Nyhoff. pp. 469-497.
Morrison, W.I. and Max Murray. 1985. The role of humoral immune responses in determining susceptibility of A/J and C57B1/6 mice to infection with Trypanosoma congolense. Parasite Immunol. 7: 63-80.
Mortelmans, J. and P. Kageruka. 1976. Trypanotolerant cattle breeds in Zaire. World Anim. Rev. 19: 14-17.
Murray, Max. 1974. The pathology of African trypanosomiasis. In: Progress in Immunology II. L. Brent and J. Holborow, eds. Amsterdam: North Holland Publishing Company, pp. 181-192.
Murray, Max and G.M. Urquhart. 1977. Immunoprophylaxis against African trypanosomiasis. In: Immunity to Blood Parasites of Animals and Man. L.H. Miller, J.A. Pino and J.J. McKelvey, Jr., eds. New York: Plenum Publishing Corporation. pp. 209-241.
Murray, Max. 1979. Anaemia of bovine African trypanosomiasis: An overview. In: Pathogenicity of Trypanosomes. G. Losos and A. Chouinard, eds. IDRC No. 132e. pp. 121-127.
Murray, Max and W.I. Morrison. 1979. Non-specific induction of increased resistance in mice to Trypanosome congolense and T. brucei by immunostimulants. Parasitology, 79: 349-366.
Murray, Max, D.J. Clifford, G. Gettinby, W.F. Snow and W.I.M. McIntyre. 1981a. A study on the susceptibility to African trypanosomiasis of N'Dama and Zebu cattle in an area of Glossina morsitans submorsitans challenge. Vet. Rec. 19: 503-510.
Murray, Max, J.G. Grootenhuis, G.W.O. Akol, D.L. Emery, S.Z. Shapiro, S.K. Moloo, Faiqa Dar, D.L. Bovell and J. Paris. 1981b. Potential application of research on African trypanosomiasis in wildlife and preliminary studies on animals exposed to tsetse infected with Trypanosoma congolense. In: Wildlife Diseases Research and Economic Development. L. Garstad, B. Nestel and M. Graham, eds. IDRC-179e. pp. 40-45.
Murray, Max, W.I. Morrison and D.D. Whitelaw. 1982. Host susceptibility to African trypanosomiasis: Trypanotolerance. Advances in Parasitology. J.R. Baker and R. Muller, eds. London: Academic Press. pp. 1-68.
Murray, P.K., Max Murray, Margaret Wallace, W.I. Morrison and W.I.M. McIntyre. 1979. Trypanosomiasis in N'Dama and Zebu cattle. II. The influence of weight of infection on the severity of the disease. International Scientific Council for Trypanosomiasis Research and Control. 15th Meeting. Banjul, The Gambia. 1977. OAU/STRC. Publication No. 110. pp. 482-487.
Njogu, A.R., R.B. Dolan, A.J. Wilson and P.D. Sayer. 1985. Trypanotolerance in East African Orma Boran cattle. Vet. Rec. 117: 632-636.
Oduye, O.O. and O.A. Okunaiya 1971. Haematological studies on the White Fulani and N'Dama breeds of cattle. Bull. Epizootiol. Dis. Afr. 19: 213-218.
Pagot, J. 1974. Les races trypanotolerance. In: Les moyens de lutte contre les trypanosomes et leurs vecteurs: actes du colloque, Paris 12-15 Mars 1974. Institut d'Elevage et de Medecine Veterinaires des Pays Tropicaux, Paris. pp. 235-248.
Paling, R.W., S.K. Moloo and J.R. Scott. The relationship between parasitaemia and anaemia in N'Dama and Zebu cattle following four sequential challenges with Glossina morsitans centralis infected with Trypanosoma congolense. International Scientific Council for Trypanosomiasis Research and Control. 19th Meeting. Lome, Togo. 1987. OAU/STRC. Publication No. 114. In press.
Pierre, C. 1906. L'elevage dans l'Afrique Occidentale Francaise. Gouvernement General de l'Afrique Occidentale Francaise, Paris.
Pinder, M. 1984. Trypanosoma congolense: genetic control of resistance to infection in mice. Exp. Parasitol. 57: 185-194.
Pinder, M., G. Libeau, W. Hirsch, I. Tamboura, R. Hauck-Bauer and G.E. Roelants. 1984. Anti-trypanosome specific immune responses in bovids of differing susceptibility to African trypanosomiasis. Immunol. 51: 247-258.
Preston, J.M. and E.W. Allonby. 1979. The influence of breed on the susceptibility of sheep to Haemonchus contortus infection in Kenya. Res. Vet. Sci. 26: 134-139.
Roberts, C.J. and A.R. Gray. 1973a. Studies on trypanosomeresistant cattle. I. The breeding and growth performance of N'Dama, Muturu and Zebu cattle maintained under the same conditions of husbandry. Trop. Anim. Hlth Prod. 5: 211-219.
Roberts, C.J. and A.R. Gray. 1973b. Studies on trypanosomeresistant cattle. II. The effect of trypanosomiasis on N'Dama, Muturu and Zebu cattle. Trop. Anim. Hlth Prod. 5: 220-233.
Roelants, G.E. 1986. Natural resistance to African trypanosomiasis. Parasite Immunol. 8: 1-10.
Rurangirwa, F.R., A.J. Musoke, V.M. Nantulya, C. Nkonge, L. Njuguna, E.Z. Mushi, L. Karstad and J. Grootenhuis. 1986. Immune effector mechanisms involved in the control of parasitaemia in Trypanosome brucei-infected wildebeest (Connochaetes taurinus). Immunol. 58: 231-237.
Saror, D.I., A.A. Ilemobade and S. Nuru. 1981. The haematology of N'Dama and Zebu cattle experimentally infected with Trypanosoma vivax. International Scientific Council for Trypanosomiasis Research and Control, 16th Meeting, Yaounde, Cameroon, 1979. pp. 287-294. OAU/STRC No. 111. Scientific Publications Division of Eleza Services, Nairobi.
Sendashonga, C.N. and S.J. Black. 1982. Humoral responses against Trypanosoma brucei variable surface antigen are induced by degenerating parasites. Parasite Immunol. 4: 245-257.
Stewart, J.L. 1937. The cattle of the Gold Coast. Vet. Rec. 49: 1289-1297.
Stewart, J.L. 1951. The West African Shorthorn cattle. Their value to Africa as trypanosomiasis resistant animals. Vet. Rec. 63: 454-457.
Toure, S.M., A. Gueye, M. Seye, M.A. Ba and A. Mane. 1978. A comparison between the pathology of a natural trypanosome infection in Zebu and N'Dama cattle. Rev. d'Elevage Med. Vet. Pays Trop. 31: 293-313.
Van Hoeve, R. 1972. Some observations on the performance of N'Dama and Muturu cattle under natural conditions in Northern Nigeria. International Scientific Council for Trypanosomiasis Research and Control. 13th Meeting. Lagos. OAU/STRC. Publication No. 105. pp. 103-106.
Vercoe, J.E. 1974. Studies on adaptation of cattle to tropical environments and the role of radioisotopes. In: Tracer Techniques in Tropical Animal Production. International Atomic Energy Agency, Vienna. pp. 83-87.
Wellde, B.T., W.T. Hockymeyer, R.M. Kovatch, M.S. Boghal and C.L. Diggs. 1981. Trypanosoma congolense: natural and acquired resistance in the bovine. Exp. Parasitol. 52: 219-232.
Whitelaw, D.D., J.A. Macaskill, P.H. Holmes, F.W. Jennings and G.M. Urquhart. 1983. Immune mechanisms in C57B1/6 mice genetically resistant to Trypanosoma congolense infection. I. Effects of immune modulation. Parasite Immunol. 5: 85-94.
![]()
![]()
![]()
{kind=link}
{kind=link}