![]()
![]()
![]()
L. V. CUNDIFF
Introduction
Additive genetic variation
Non-additive genetic variation
Conclusion
References
Genetic resistance to disease is undoubtedly under natural selection as a strong component of fitness. Resistance to disease may have been increased indirectly through favourable associations with other characters such as growth rate. However, except for some selection against mastitis incidence in dairy cattle (Lindhe, 1982; Miller, 1982), genetic resistance to disease has not received direct emphasis in genetic improvement programmes for cattle. Increased research effort on genetics of disease resistance in cattle is justified even though the economics of beef or dairy production are dominated by other traits. Crossbreeding systems are available which can exploit heterosis and the vast genetic variation that exists among and within breeds for retail product growth, milk production and other economically important biological characteristics to match genetic potential with climate, feed resources, and market requirements (Gregory and Cundiff, 1980; Cundiff et al., 1986). Nearly optimal levels of performance for traits such as retail product growth, milk production, age at puberty and mature size can be achieved with relative ease by use of properly chosen breeds in appropriate crossbreeding systems. Once optimal or nearly optimal levels of production for these important traits are achieved, more attention can be focussed on other characteristics such as reproduction, survivability, and genetic resistance to disease which have received less attention because of low heritability and complications of diagnosis and recording.
The purpose of this paper is to review quantitative genetic approaches to breeding for genetic resistance in cattle. The classical quantitative genetics approach to breeding for genetic improvement of any characteristic is to first assess and then utilize 1) additive genetic variation between breeds, 2) additive genetic variation within breeds, 3) non-additive genetic variation between breeds (heterosis), and 4) non-additive genetic variation within breeds (inbreeding). Procedures for experimental assessment and utilization of each of these sources of genetic variation will be discussed in this review.
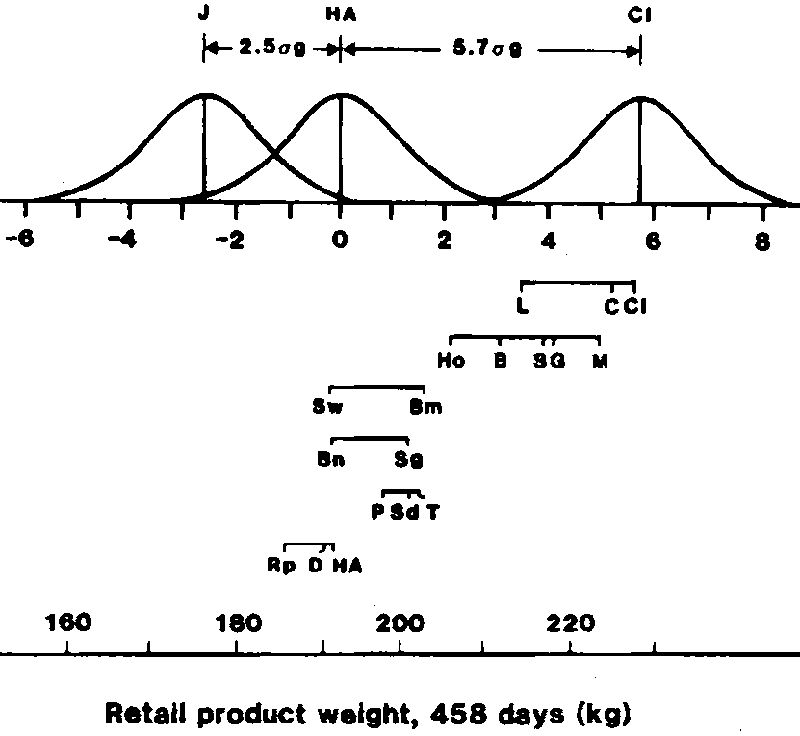
The additive genetic variation that exists among and within breeds of cattle for many production characteristics is vast. Results from the Germ Plasm Evaluation Program at the Roman L. Hruska U.S. Meat Animal Research Center are shown in Figure 1 for one important measure of output, retail product weight (closely trimmed-boneless steaks, roasts and lean trim). In Figure 1, F1 cross means for weight of retail product at 458 days of age are shown on the lower horizontal axis. The spacing on the vertical axis is arbitrary, but the ranking from the bottom to top reflects increasing increments of mature size. Steers sired by bulls of breeds with large mature size produced significantly more retail product weight than steers sired by breeds of small mature size. Differences are doubled in the upper horizontal scale to reflect variation among pure breeds relative to a standard deviation change in breeding value (s g=(s 2p) (h2)) within pure breeds. Frequency curves, shown for Jersey, the average of Hereford and Angus, and Chianina, reflect the distribution expected for breeding values of individual animals within pure breeds. The breeding value of the heaviest Jersey is not expected to equal that of the lightest Chianina and the heaviest Hereford and Angus would only equal the lightest Chianina in genetic potential for retail product growth to 458 days. The range between breeds (8.2s g) exceeded that expected for breeding value of individuals within breeds (6s g). Both between and within breed sources of genetic variation were large and important for this measure of output-retail product weight. Similar results have been found for other biological characteristics important to beef production.
Figure 1. Variation between and within breeds.
Historically, we have often locked in on the use of a few or even one breed because of their excellence in a single trait (e.g. milk production) and overlooked opportunities to exploit genetic variance among breeds for other characteristics. Genetic variance between breeds for resistance to disease should be considered. Heritability of breed differences is very high, provided the breed means are estimated with an adequate sample to average out errors of sampling individual animals within breeds. The tendency for progeny from individual animals to regress to their own breed group mean is much greater than any tendency to regress to the mean of all cattle. However, because mean differences between breeds are so highly heritable, once between-breed genetic variation has been exploited by selection of the desired breed or breeds used in a crossbreeding programme or in foundation of a composite population, continued genetic improvement is dependent on intrapopulation selection and genetic variation available within the breeds or composite population. Intrapopulation (i.e., within-breed or composite) variation is virtually restored generation after generation by the Mendelian process, while variation between populations, accruing only very slowly as a result of different selection goals, genetic drift (associated with inbreeding) or rare mutations, can only be exploited rarely (e.g. at times abrupt changes in selection goals are indicated and exotic germ plasm is available). Although intrapopulation genetic variation should have the greatest long-term impact on genetic change for any specific objective, opportunities to optimize performance levels by selection among breeds should not be overlooked.
Additive genetic variation among breeds
Additive genetic variation among breeds can be assessed in experiments involving breed comparisons. Diallel crossing studies are most informative because they provide for estimation of direct and maternal effects and enable estimation of non-additive gene effects as well, but they are not as practical for evaluation of a large number of breeds as topcross comparisons involving females of one, or more prevalent breeds. Unfortunately, disease status of animals has been reported in only a limited number of breed comparison studies.
Significant genetic variation has been shown among breeds of cattle for genetic resistance to certain pathogens and parasites. The N'Dama and West African Shorthorn are significantly more resistant to trypanosomiasis than Bos indicus breeds (Murray and Trail, 1982). Recent evidence indicates that a relatively high proportion of Baoule cattle are more resistant than Bos indicus cattle (69% resistant, 31% as sensitive as Bos indicus to trypanosomiasis (Roelants et al., 1987). Indications are that Bos indicus breeds are intermediate in resistance to trypanosomiasis to the N'Dama and West African Shorthorn and Bos taurus breeds from Europe.
Bos indicus cattle and ii x Bos taurus crosses are significantly more resistant to ticks than Bos taurus breeds or Bos taurus x Bos taurus breed crosses. In addition, Frisch and Vercoe (1978) have shown that Bos indicus x Bos taurus breed crosses (1/2 Brahman, 1/4 Hereford, 1/4 Shorthorn; and 1/2 Africander, 1/4 Hereford, 1/4 Shorthorn) are significantly more resistant to gastrointestinal helminths and pink-eye disease (bovine keratoconjunctivitis) than Bos taurus x Bos taurus crosses (Hereford x Shorthorn).
For immunity to many pathogens, young calves are dependent on immunoglobulins passively received from their dam in colostrum. Significant differences were found between Angus, Red Poll and Herefords in ability to acquire and absorb colostral antibodies (Muggli et al., 1984). Breeds ranked Angus (39 mg/ml), Red Poll (35 mg/ml) and Hereford (30 mg/ml) for Ig G1 concentration.
Herefords are significantly more susceptible to cancer eye than Angus and Shorthorns or F1 crosses of these breeds (Nunez-Domenquez et al., 1984). It is likely that significant variation exists among breeds for resistance to other pathogens; however, further studies are needed to assess genetic variation among breeds for general disease resistance and for resistance to specific pathogens. Disease status of animals needs to be monitored more closely in breed comparison studies.
Additive genetic variation within breeds
Additive genetic variation within breeds is ordinarily expressed as a function of heritability (h2) and phenotypic variation (s 2p),
s 2g = h2s 2p
Heritability is estimated from the extra likeness of relatives compared to likeness of non-relatives (e.g., offspring on parent regression, paternal half-sib analysis of variance, or full-sib analysis of variance). Realized heritability can be estimated directly from long-term single trait selection experiments.
Heritability of overall fitness traits such as survival are low. Heritability tends to be higher when treated as a trait of the dam (8%) than when treated as a trait of the offspring (4%), (Cundiff et al., 1982). When the large number of causes of mortality are considered, it is not surprising that heritability of survival is low. However, specific components of survival have moderate to high heritability.
Mastitis
Resistance to mastitis is 10 to 20% heritable (Miller, 1982). Attention is given to mastitis susceptibility in cattle genetic improvement programmes in Sweden (Lindhe, 1982) and Norway (Solbu et al., 1982). A possible association between bovine major histocompatibility (BoLA) genotype and mastitis susceptibility has been shown by Solbu et al. (1982). A significant association was found between BoLA genotype determined for 130 young bulls and breeding values of their sires for mastitis susceptibility (based on progeny tests of first lactation daughters). One BoLA type (W2) was associated with high resistance to mastitis while another (W16) was associated with susceptibility to mastitis.
Tick resistance
Resistance to ticks has been found to be highly heritable and responsive to selection (Utech and Wharton, 1982). After just three generations of selection, resistance increased from 89 to 99% in an Illawara Shorthorn herd. In a Brahman x Australian Illawara Shorthorn population, resistance to ticks increased from 97.6 to 99.6% in response to three generations of selection. Resistance to ticks was determined from the average percent mortality of female ticks from two artificial infestations with 20,000 larvae, 14 days apart.
Intense population screening, to identify animals with extreme or rare phenotypes (Gregory et al., unpublished; Piper and Bindon, pers. comm.) followed by a multiplication aided by use of superovulation and embryo transfer has accelerated the rate of progress at least in the first generation of selection for traits that are expressed rarely such as twinning (Gregory et al., unpublished; Piper and Bindon, pers. comm.). It would be interesting to try this technique among animals that have survived epidemic challenges to specific diseases such as trypanosomiasis. Such a procedure should capture genetically resistant animals regardless of whether or not their resistance is attributable to many genes with small effects, a few genes with large effects or many genes some of which have large effects. Identification of major genes should be facilitated in experiments which can contrast unselected to intensively screened and selected animals and progeny.
Major genes associated with specific diseases
Nicholas (1987) reviewed evidence of associations between the MHC and specific diseases. One experiment in poultry demonstrated how high versus low selection for specific diseases can facilitate identification of major genes associated with genetic resistance to disease. In this experiment, selection for resistance versus susceptibility to Marek's disease, a neoplastic disease in chickens caused by a DNA virus, was compared. Incidence of the disease was 51% in the foundation stock. After four generations of selection, mortality in those selected for resistance was 7% compared to 94% in those selected for susceptibility. Early indications were that susceptibility was associated with an MHC allele (B19) and resistance was associated with an MHC allele (B21). Subsequent experimentation confirmed that the B21 allele was indeed associated with resistance, while B19 and other alleles at the B locus are associated with susceptibility to Marek's disease.
In cattle, associations have been reported between BoLA class I antigens and resistance to ticks, worm infestation, ocular squamous carcinoma and bovine leucosis (Steer et al., 1984).
General disease resistance
There are many examples of successful selection for resistance to specific diseases in mice and poultry (e.g., Gavora and Spencer, 1983). However, as Gavora and Spencer (1983) have pointed out, successful selection for specific resistance to all diseases of animals is impossible, especially if appropriate attention is to be given to other important production characters. Thus, they proposed that selection criteria be developed for general disease resistance. The most encouraging evidence in support of this approach, at least for certain groups of diseases, comes from selection studies for immunoresponsiveness which have been conducted in mice (Biozzi et al., 1982).
Table 1. Characteristics of five selections for high (H) vs low (L) antibody production in mice (Biozzi et al., 1982)
|
Sell |
Antigens used proc. |
Immunization |
No. gent to sel. limit |
Diff. (H/L) |
h2 |
No. of independ. loci |
|
I |
Sheep eryth. |
Primary |
16 |
220 fold |
.20 |
9-11 |
|
|
Pigeon eryth. |
Response |
|
|
|
|
|
II |
Sheep eryth. |
Primary |
13 |
103 fold |
.21 |
2-8 |
|
|
response |
|
|
|
|
|
|
III |
Salm. typh. |
Secondary |
16 |
90 fold |
.20 |
4-7 |
|
|
Salm. van. |
response |
|
|
|
|
|
IV |
Salm. typh. |
Secondary |
12 |
85 fold |
.21 |
2-4 |
|
|
Salm. van. |
response |
|
|
|
|
|
V |
Bov. Ser. Alb. |
Hyperimmun |
7 |
310 fold |
.22 |
2-4 |
|
|
Rabbit gamma. glob |
alum precip. antigen |
|
|
|
|
In their work, mice were selected for high versus low peak antibody titers after optimal immunization using five different regimens involving a variety of complex natural immunogens (Table 1). After a plateau of selection responses when differences between the high and low lines reached a maximum value, the lines were assumed homozygous for loci regulating the characters under selection. All selections diverged significantly, and remarkably consistent realized heritabilities were noted. The moderately high heritability estimates indicate that significant additive genetic variation does exist for specific components of resistance to disease. The range of number of loci estimated by different methods provides evidence that antibody production is subject to polygenic regulation. Comparison of F2 hybrids homozygous for the H-2 phenotype of parental H and L lines indicated that the major histocompatibility complex accounted for only part (14 to 26%) of the interline difference produced by the additive effect of all loci involved.
Resistance to a variety of bacterial and parasitic infections has been studied in Selection I (Table 2). The high response line was more resistant to infections dependent upon antibody immunity, while the low line was more resistant to infections dependent upon microphage immunity. In most cases, the line that was spontaneously more resistant also was protected by vaccination to a higher degree.
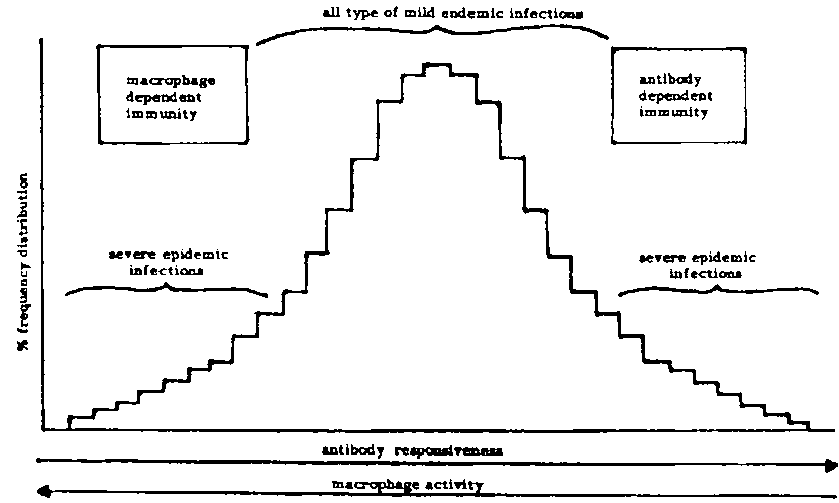
On the basis of their results, Biozzi et al. (1982) have proposed a theory for the evolution of host-parasite interactions in genetically heterogeneous populations (Figure 2). They proposed that F2 hybrids between homozygous H and L mice would present a normal frequency distribution of phenotypes, similar to that which is expected in genetically heterogeneous natural animal populations. The axes would have inversed directions for antibody production and for macrophage activity. The tails of the distribution would be represented by individuals that were extremely resistant or highly susceptible to severe epidemic infections depending upon which defence mechanism is responsible for protection against the infection. Thus, at least a few individuals would survive any sudden severe epidemic infection. However, the majority of individuals falling somewhere between the two extremes would possess a median resistance to disease, being protected somewhat by both antibody production and macrophage activity. It was proposed that this type of genetic polymorphism would be maintained by stabilizing natural selection under conditions of mild endemic infection when extreme individuals at the distribution tails would be eliminated. Such a model would be consistent with the many polymorphisms found in human populations (e.g., Brock and Mayo, 1978) and with significant effects of heterosis on survival, longevity, resistance to ticks and other stresses in cattle and other species. Mating systems that maximize heterozygosity would lead to increased genetic resistance to disease under this model of overdominance for survival.
Table 2. Summary of results on resistance of H and L mice to various infections (Biozzi et al., 1982)
|
|
Infection |
Degree of resistance |
||||
|
Innate resistance |
Acquired resistance |
|||||
|
|
|
H |
L |
H |
L |
|
|
P. berghei |
- |
- |
+++ |
± |
||
|
T. cruzi |
+++ |
- |
n.d. |
- |
||
|
Antibody |
||||||
|
|
dependent |
N. dubius |
- |
- |
+++ |
± |
|
|
immunity |
Rabies virus* |
- |
- |
+++ |
± |
|
|
T. spiralis |
++ |
± |
++ |
++ |
|
|
S. typhimurium |
± |
++ |
+ |
++++ |
||
|
Macrophage |
Y. pestis |
± |
++ |
+ |
++++ |
|
|
|
dependent |
B. abortus suds |
+ |
++ |
++ |
++++ |
|
|
immunity |
L. tropica |
+ |
+++ |
n.d. |
n.d. |
|
|
S. mansoni |
+ |
++ |
+++ |
++ |
|
*Experiments carried out in Selection III and IV.
Non-additive genetic variation between breeds
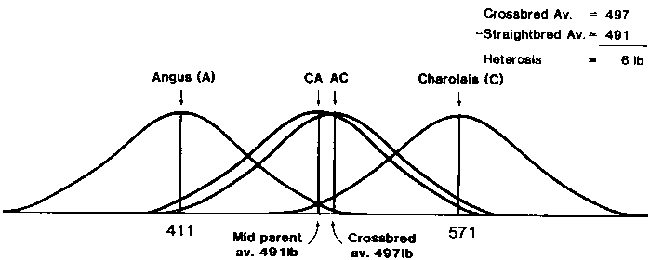
Non-additive genetic effects among breeds are assessed by evaluating effects of heterosis in breed crosses. Effects of heterosis are estimated from the difference between the mean for reciprocal FL crosses and the mean for parental purebreds in diallel crossing experiments. It is important to evaluate reciprocal F1 crosses to avoid confounding of heterosis with additive genetic maternal effects. For highly heritable traits, such as retail product, effects of heterosis are relatively small and the mean of F1 crosses is about intermediate to that of parental purebreds (Figure 3). On the other hand effects of heterosis are often rather large relative to genetic differences between breeds for traits that are lowly heritable. Unfortunately, disease incidence has not been reported in diallel crossing experiments with cattle. Significant effects of heterosis have been demonstrated for components of fitness including reproduction
Figure 3. Breed group values for retail product.
Survival
Effects of heterosis on survival are large and important. Calf crop percentages weaned are increased by 3.4% by effects of individual heterosis on survival and by an additional 1.3% by effects of maternal heterosis on survival of calves raised by F1 cross dams (Long, 1980). These results indicate that significant differences have evolved between breeds in frequency of genes that have non-additive effects (e.g. dominance) on survival of calves. The generally consistent estimates of heterosis among experiments and among years suggests that resistance to a number of stresses associated with mortality is greater in F1 crosses than in parental purebreds. Evidently, increased heterozygosity at a number of loci are involved.
Table 3. Breed group means (years) for longevity of crossbred and straightbred cows (Nunez et al., 1984)
|
Breed of dam of cow |
Breed of sire of cow |
||||
|
Hereford |
Angus |
Shorthorn |
Average |
||
|
Hereford |
8.46 |
11.04 |
9.61 |
9.70 |
|
|
Angus |
10.60 |
9.41 |
9.90 |
9.97 |
|
|
Shorthorn |
8.01 |
9.26 |
7.28 |
8.18 |
|
|
|
Average |
9.02 |
9.90 |
8.93 |
9.28 |
|
|
Crossbred |
Straightbred |
Difference |
||
|
Heterosis |
9.74 |
8.38 |
1.36 (16.2%) |
||
Longevity
Significant differences have been found among breeds and breed crosses in longevity (Nunez-Domenquez et al, 1984). Angus had significantly greater longevity than Shorthorns (Table 3). Effects of heterosis increased average longevity of F1 cows by 1.36 years over that of straightbred Hereford, Angus, and Shorthorn cows. Reasons for removal and average age at removal of crossbred and straightbred cows are summarized in Table 4, some of which involve differences in genetic resistance to disease. The four straightbred cows, removed because of cancer eye, were all Herefords. Cancer eye susceptibility is known to be a highly heritable component of longevity (Anderson et al., 1957a; 1957b). Death losses were significantly greater in straightbreds than crossbreds. Unsound udders developed more frequently in crossbred cows than in straightbred cows, but problems developed at older ages in crossbreds than in straightbreds. About 19% of the crossbreds compared to 6.5% of the straightbreds remained in the herd from initiation to termination of the experiment when the cows ranged from 12 to 15 years of age. Significant effects of heterosis on longevity have also been reported by Spelbring et al. (1977).
Non-additive genetic variation within breeds
Non-additive genetic variation within breeds is assessed by evaluating effects of inbreeding within inbred lines and by estimating effects of heterosis in diallel crossing experiments involving lines. Effects of inbreeding depression are significant for fitness traits (Brinks et al. 1975).
Table 4. Reasons for and average age at removal from cow herd (unpublished).
|
Reason |
Straightbred |
Crossbred |
|||||
|
No. |
% |
Age, yr |
No. |
% |
Age, yr |
||
|
Death |
31 |
19.9 |
10.1 |
18 |
10.4 |
9.0 |
|
|
Open |
85 |
54.5 |
6.5 |
91 |
52.8 |
7.8 |
|
|
Crippled |
5 |
3.2 |
10.6 |
7 |
4.0 |
9.5 |
|
|
Unsound udder |
0 |
0.0 |
- |
9 |
5.3 |
13.0 |
|
|
Cancer eye |
4 |
2.5 |
11.1 |
0 |
0.0 |
- |
|
|
Lump jaw |
1 |
.6 |
8.6 |
0 |
0.0 |
- |
|
|
Prolapse |
3 |
1.9 |
4.3 |
0 |
0.0 |
- |
|
|
Emaciation |
11 |
7.1 |
12.0 |
7 |
4.0 |
13.1 |
|
|
Unknown |
6 |
3.8 |
5.4 |
7 |
4.1 |
4.6 |
|
|
Exp. termin. |
10 |
6.5 |
13.1 |
33 |
19.4 |
13.2 |
|
|
|
Total or average |
156 |
100.0 |
8.4 |
172 |
100.0 |
9.7 |
Significant additive Genetic variation exists among breeds for survival, longevity, and resistance to tropical environmental factors (i.e. trypanosomiasis, ticks, gastrointestinal helminths, high ambient temperature, solar radiation, pink-eye disease and nutritional stress). Heritability of survival (dead or alive) is low, but specific components of survival and longevity are moderately to highly heritable (e.g., packed cell volume/trypanotolerance, resistance to ticks, immunoglobulin concentration, mastitis, bloat, and cancer eye). Studies with mice indicate that selection for immunoresponsiveness to specific infectious agents may lead to increased resistance to a certain group of infections but increased susceptibility to other groups of infections depending upon the protective mechanism involved. To manage antagonistic genetic relationships such as these, it may be necessary to select for divergence between paternal and maternal strains (or breeds), which when crossed, complement each other to achieve optimal genetic resistance to disease and adaptation to the climatic and nutritive environment. Non-additive genetic effects (heterosis) are important for survival and longevity.
Anderson, D.E., J.L. Lush and D. Chambers. 1957a. Studies on bovine ocular squamous carcinoma (cancer eye). II. Relationship between eyelid pigmentation and occurrence of cancer eye lesions. J. Anim. Sci. 16:739.
Anderson, D.E., D. Chambers and J.L. Lush. 1957b. Studies on bovine ocular squamous corcinoma (cancer eye). III. Inheritance of eyelid pigmentation. J. Anim. sci. 16:1007.
Biozzi, G., D. Mouton, A.M. Heumann and Y. Bouthillier. 1982. Genetic regulation of immunoresponsiveness in relation to resistance against infectious diseases. Proceedings of the 2nd World Congress on Genetics Applied to Livestock Production. Vol. V. Madrid. pp. 150-163.
Brinks, J.S. and B.W. Knapp. 1975. Effects of Inbreeding on Performance of Beef Cattle in the Western Region. A Western Regional Publication. Colorado State U., Fort Collins, Colorado.
Brock, D.J.H. and O. Mayo. 1978. The Biochemical Genetics of Man. 2nd edition. London: Academic Press.
Cundiff, L.V., K.E. Gregory and R.M. Roch. 1982. Selection for increased survival from birth to weaning. Proceedings of the 2nd World Congress on Genetics Applied to Livestock Production. Vol. V. pp. 310-337.
Cundiff, L.V., K.E. Gregory, R.M. Koch and G.E. Dickerson. 1986. Genetic diversity among cattle breeds and its use to increase beef production efficiency in a temperate environment. Proceedings of the 3rd World Congress on Genetics Applied to Livestock Production. Vol. IX: 271-282.
Frisch, J.E. and J.E. Vercoe. 1978. Utilizing breed differences in growth of cattle in the tropics. Wld Anim. Rev. 25:8-12.
Gavora, J.J. and J.L. Spencer. 1983. Breeding from immune responsiveness and disease resistance. Anim. Blood Groups and Biochem. Genetics 14:159-180.
Gregory, K.E. and L.V. Cundiff. 1980. Crossbreeding in beef cattle: Evaluation of systems. J. Anim. Sci. 51:1224.
Lindhe, B. 1982. Possibilities to include disease resistance in dairy cattle breeding. Proceedings of 2nd World Congress on Genetics Applied to Livestock Production. Vol. VII. pp. 336343.
Long, C.R. 1980. Crossbreeding for beef production. Experimental results. J. Anim. Sci. 51: 1197.
Miller, R. 1982. Genetics of resistance to mastitis. Proceedings of the 2nd World Congress on Genetics Applied to Livestock Production, Madrid. Vol. V. pp. 186-198.
Muggli, N.E., W.D, Hohenboken, L.V. Cundiff, and K.W. Kelley. 1984. Inheritance of maternal immunoglobulin G1 concentration by the bovine neonate. J. Anim. Sci. 59: 39-48.
Murray, M. and J.C.M. Trail. 1982. Trypanotolerance: genetics, environmental influences and mechanisms. Proceedings of the 2nd World Congress on Genetics Applied to Livestock Production. Vol. VI: 293-306.
Nicholas, F.W. 1987. Veterinary Genetics. Oxford: Clarendon Press.
Nunez-Domenquez, R.D., L.V. Cundiff, G.E. Dickerson and K.E. Gregory. 1984. Heterosis of longevity in beef cattle. J. Anim. Sci. Abstracts. 59: (Suppl. 1) 180-181.
Roelants, G.E., F. Fumoux, M. Pinder, R. Querol, A. Bassinga and E. Authie. 1987. Identification and selection of cattle naturally resistant to African trypanosomiasis. Acta Tropica 44: 55-66.
Spelbring, M.C., T.G. Martin and K.J. Drewry. 1977. Maternal productivity of crossbred Angus x Milking Shorthorn cows. II. Cow productivity and longevity. J. Anim. Sci. 45:976.
Solbu, H., R.L. Spooner and O. Lie. 1982. A possible influence of the bovine major histocompatibility complex (BoLA) on mastitis. Proceedings of the 2nd World Congress on Genetics Applied to Livestock Production. Vol. VII. Madrid. pp. 368-371.
Stear, M.J., M.J. Newman, F.W. Nicholas, S.C. Brown and R.G. Holmoyd. 1984. Aust. J. Exp. Biol. Med. Sci. 62:47-52.
Utech, K.B.W. and R.H. Wharton. 1982. Breeding for resistance to Boophilus microplus in Australian Illawarra Shorthorn and Brahman x Australian Illawarra Shorthorn cattle. Aust. Vet. J. 58:41-46.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}