![]()
![]()
![]()
by
B. IVANOV
All-Union Research Institute
of Marine Fisheries and Oceanography
Moscow, U.S.S.R.
Abstract
Explorations in the North Pacific in recent years have revealed considerable stocks of Pandalus borealis Krøyer in the Bering Sea and the Gulf of Alaska. Studies on size composition show that the shrimp catches are mostly made up of specimens aged between 1½ and 5½ years; a few survive to 6½ years. Size at first maturity (as males), at sex-change and at second maturity (as females) was also studied. The strength of different year-classes can be largely explained from the character of the first winter after hatching and the abundance of the parent year-classes.
Three ecological zones are recognizable within the area considered. Large concentrations of shrimps tend to form in the zone of interaction of two water masses rather than in waters of a particular temperature. Local circulations due to islands and bays help to prevent the dispersal of larvae to unfavourable areas.
BIOLOGIE ET REPARTITION DE LA CREVETTE NORDIQUE DANS LA MER DE BERING ET LE GOLFE DE L'ALASKA
Résumé
Des campagnes de prospection menées dans le Pacifique Nord ces dernières années ont révélé la présence de stocks considérables de Pandalus borealis Krøyer dans la mer de Béring et le golfe de l'Alaska. L'étude de la composition par taille a indiqué que les captures comprennent principalement des crevettes d'un âge situé entre 1,5 et 5,5 ans; quelques unes atteignent 6,5 ans. Les recherches ont porté également sur la taille à la première maturité (en tant que mâles), à l'inversion du sexe et à la seconde maturité (en tant que femelles). L'importance des différentes classes d'âge s'explique en grande partie par les caractéristiques du premier hiver qui suit l'incubation et par l'abondance de la classe d'âge des géniteurs.
On distingue trois zones écologiques dans la région étudiée. Le fortes concentrations de crevettes tendent à se constituer plutôt dans la zone d'interaction de deux masses d'eau que dans des eaux d'une température déterminée. La présence d'îles et de baies détermine des circulations locales qui contribuent à empêcher la dispersion des larves vers des régions défavorables.
LA BIOLOGIA Y DISTRIBUCION DEL CAMARON SEPTENTRIONAL EN EL MAR DE BEHRING Y EN EL GOLFO DE ALASKA
Extracto
Las exploraciones efectuadas en el Pacífico septentrional durante estos últimos años han revelado la existencia de considerables poblaciones de Pandalus borealis Krøyer en el Mar de Behring y en el Golfo de Alaska. Los estudios sobre la composición por tallas indican que las capturas de camarones están formadas en su mayor parte por ejemplares de edades comprendidas entre 1,5 y 5,5 años; algunos sobreviven hasta los 6,5 años. También se estudió la talla durante la primera madurez (en los machos) durante el cambio de sexo y en la segunda madurez (en las hembras). El volumen de las diferentes clases de edad se puede explicar, en gran parte, por las características del primer invierno después de la eclosión y por la abundancia de la clase de edad de los padres.
Dentro del área considerada se pueden reconocer tres zonas ecológicas. En la zona de interacción de dos mases de agua tienden a formarse grandes concentraciones de camarones en vez de formarse en aguas de una determinada temperatura. Los movimientos de agua de tipo local que tienen lugar, debido a la existencia de islas y bahías, contribuyen a evitar la dispersión de las larvas hacia zonas desfavorables.
The many exploratory cruises conducted in the last few years in the North Pacific have revealed appreciable stocks of shrimp in the Bering Sea and the Gulf of Alaska. Now the stocks are exploited by the fishermen of Japan, the U.S.A. and U.S.S.R. The present paper is a brief summary of the Soviet investigations on the biology and distribution of the Pandalus borealis Krøyer, a species of a great commercial interest in the Bering Sea and the Gulf of Alaska. The carapace length (cl), from the posterior limit of the eye socket to the extreme posterior lateral edge of the carapace, is the main size measurement used.
2.1 The Bering Sea
Data for the Pribilof area of the Bering Sea are available for August, November and December 1962; January, February, August 1963; May 1964; and May 1965. The shrimp size composition in trawl catches for those months is given in Fig. 1. The growth of the shrimps in the Pribilof area is represented in Fig. 2. From the diagrams of the shrimp size and sex composition it can be seen that in the Pribilof area shrimps at the age of 1½yr (cl = 12 to 13 mm) are still not sexually mature. Some shrimps at the age of 2½ yr (cl = 18 to 19 mm) still remain immature, while others become males and participate for the first time in the autumn breeding. Shrimps 3½ yr old (cl = 22 mm) are nearly all males, and only a small minority are females. Most specimens of 4½ yr (cl = 24 to 25 mm) again act as males, and only at the age of 5 yr do the bulk of shrimps change their sex. At the age of 5½ yr (cl = 27 mm), in autumn, they begin to spawn. A very small number of shrimps can survive up to 6½ yr, and many of these are sterile females. Thus, in the Pribilof area the majority of females have only one brood.
From the peaks in the diagrams of the size composition one can estimate the relative strength of shrimp year-classes. If all the age groups of shrimps (that is shrimps at the age of 1½, 2½, 3½, 4½ and 5½ years) had been equally represented in the catches, the shrimps with cl = 12 to 13, 18 to 19, 21 to 22, 24 to 25 and 27 mm should have formed the peaks in the August diagrams of size composition. But, the peaks are different sizes and can be even practically absent (Fig. 1). (Shrimps in their first year are poorly represented in the samples and are omitted from the figure). For instance, in August 1962 shrimps with cl = 18 to 19 mm formed no noticeable peak in the catches, but shrimps of the same size showed a well marked peak in August 1963. Shrimps with cl = 24 mm also did not form an obvious peak in August 1962, but they did in August 1963. As seen from 1962 and 1963 diagrams, the 1960 year-class was extremely poor, and was practically absent, the 1959 year-class was rich, the 1958 year-class was average, the 1957 year-class was rich, and the 1961 year-class was, apparently, rich.
Comparing the strength of the 1957–61 year-classes with ice conditions in the Bering Sea (Fig. 3) one can notice that year-classes which went through a mild or moderate winter in the first year of life can usually be considered as rich, and the shrimps representing the poor 1960 year-class were hatched before a severe winter. Thus, fluctuations in the abundance of the Pribilof shrimps can be fairly well explained by the assumption that the strength of year-class depends on the character of the first winter after hatching.
This assumption and records of ice conditions in the Bering Sea in the previous years (Kryndin, 1964) make it possible to estimate the strength of the parent year-classes which gave rise to the 1957–61 broods (Fig. 3). We assume that all the females in the hatching period were 6 yr old. Assuming that the abundance of the year-classes depends to some extent on the strength of their parent year-classes, we can explain further the fluctuations in the abundance of the 1957–61 year-classes.
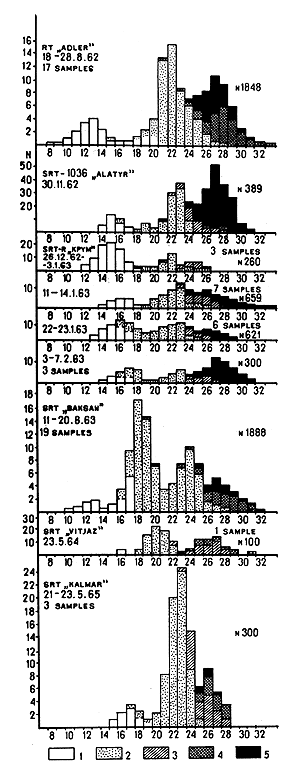
Fig. 1 Size and sex composition in P. borealis
catches in the Bering Sea (axis X, carapace length):
1 Juvenile, 2 Males, 3, Transitional
4 Females, 5 Ovigerous females
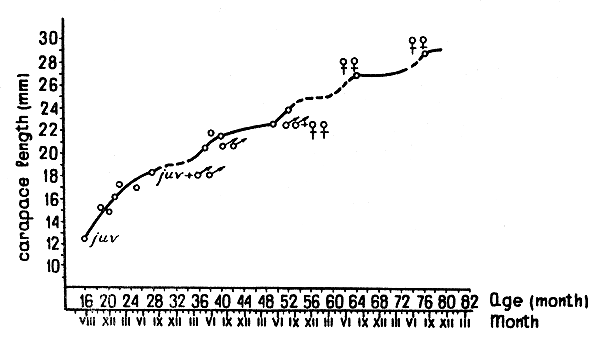
Fig. 2 Growth of P. borealis in the Bering Sea.
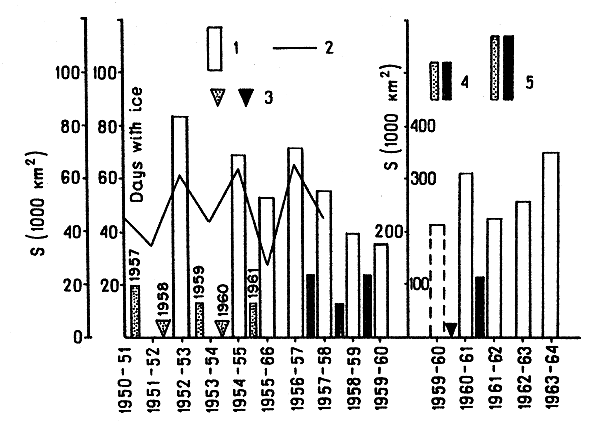
Fig. 3 Relation between ice conditions in winter and the strength of the P. borealis year-classes in the Bering Sea:
1 Sea area covered by ice in thousands of km2 (left, western Bering Sea (after Kryndin, 1964); right, whole of Bering Sea)
2 Days with ice (after Kryndin, 1964)
3 Poor year-class
4 Average year-class
5 Rich year-class (dotted, parent year-classes; black, the considered year-classes)
The fact that the 1958 generation was of only moderate strength, in spite of the mild winter which followed its hatching, can be explained by the supposed low abundance of the parent year-class of 1952.
Interrelations between the strength of year-classes, severity of the winter in the first year of shrimp life, the supposed strength of the parent year-classes are given in Table I. So, it is possible that the strength of different year-classes of the Pribilof shrimps depends on the winter character in the first year of the shrimp life and to a lesser degree on the abundance of the parent year-classes.
TABLE I
The relation between the strength of year-classes, the winter climate during the first year of shrimp life and the supposed strength of parent year-classes
| Year-class | Birth of parent year-classes | Winter character in the first yr of life of parent year-classes | Supposed strength of parent year-class | Winter character in the first yr of life of the given year-class | Strength of the year-class |
| 1957 | 1951 | mild | rich | moderate | rich |
| 1958 | 1952 | severe | poor | mild | average |
| 1959 | 1953 | moderate | average | mild | rich |
| 1960 | 1954 | severe | poor | severe | very poor |
| 1961 | 1955 | moderate | average or rich | mild | rich |
Taking into account the short series of our observations, we consider this as a preliminary conclusion to be checked by further investigations.
Since the Japanese fishermen carry out an intensive fishery for the Pribilof shrimps, it is possible that data on the catches of the various year-classes will help to check our conclusions in the near future.
2.2 The Western Part of the Gulf of Alaska
Data for the west part of the Gulf of Alaska (Kodiak-Sanak Islands) are available for December 1962, August 1963, March and May 1965 (Fig. 4). The growth of shrimps in the west part of the Gulf of Alaska is represented in Fig. 5. Because of the lack of continuous series of observations it is difficult to assess growth, sex maturation and fluctuations in the abundance of different year-classes.
It seems that in the western Gulf of Alaska shrimps at the age of 1½ yr (cl = 13 mm) remain immature and first participate in breeding as males at 2½ yr (cl = 19 mm).
Most specimens of 3½ yr (cl = 22 mm) again act as males, but some of them change sex and spawn. Change of sex takes place, in the main, at the age of 4 yr, and the majority of shrimps spawn for the first time at the age of 4½ yr (cl = 25 mm). Unlike the Pribilof shrimp population, in the Gulf of Alaska numerous females breed twice, as 4½ and 5½ yr old specimens (cl = 27 to 28 mm). Very few specimens live till 6½ yr (cl = 30 mm). Thus, in the western Gulf of Alaska sex development takes place one year earlier than in the Bering Sea.
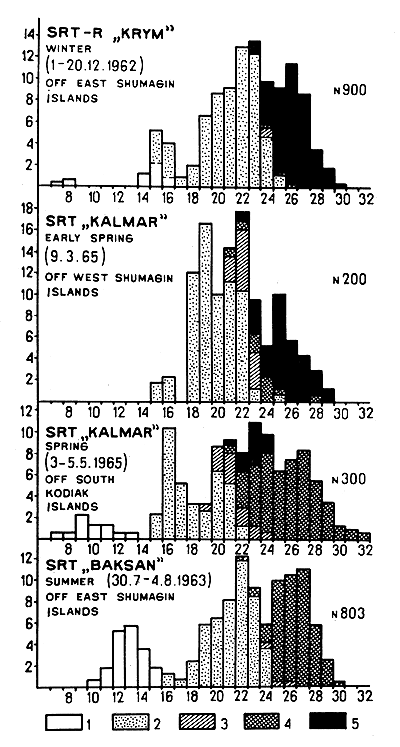
Fig. 4 The size and sex composition in P. borealis catches in the western part of the Gulf of Alaska. (Signs used are the same as for Fig. 1)
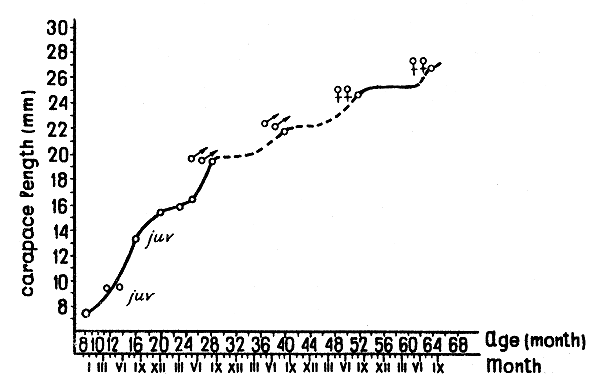
Fig. 5 The growth of the P. borealis in the western Gulf of Alaska.
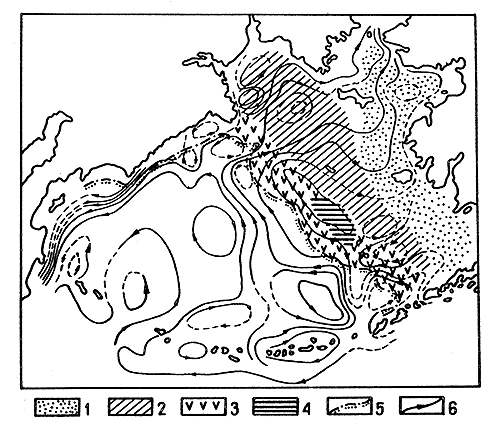
Fig. 6 Map of surface currents in the Bering Sea in June (after Arsenyev and Shcherbinin, 1963) with the location of the three ecological zones and the area of the Pribilof shrimp concentration :
As seen from the diagrams (Fig. 4), in the Gulf of Alaska (unlike the Bering Sea) sharp fluctuations in the strength of different year-classes are not observed. This fact can be, apparently, explained by the milder conditions. Thus, taking into consideration that a number of Alaskan females spawn twice, the lack of great fluctuations in the year-class strength and the more rapid sexual development of shrimps as compared with the Bering Sea, one can conclude that the western Gulf shrimp concentrations are more stable than those in the Bering Sea.
The western part of the Gulf of Alaska contrasts with the eastern part (Butler, 1964; Anon., 1965), and with European waters (Rasmussen, 1953; Horsted and Smidt, 1956; Allen, 1959), in that growth and sexual development of shrimps do not slow down towards the north. For instance, sex change of shrimps in the Shumagin Islands takes place at 4 yr, the same age as in the northern Gulf (5° further north) Anon., 1965. There is even a slight tendency for growth and sexual development to be faster in the northern than in the south-western Gulf. The size and sex composition of shrimp populations in the western Gulf of Alaska are relatively uniform over a very wide range of both latitude and longitude, and correlations with latitude may even be reversed from those found on the Norwegian coast. Comparable conditions are not found elsewhere within the range of P. borealis, but they are explained by the warm current which flows through the western Gulf from northeast to southwest. This results in milder hydrological conditions in the northern part of the area than in the southwest.
3.1 The Bering Sea
Three ecological zones, which are mainly distinguished by temperature, can be recognized on the Bering Sea shelf. The zones stretch like belts parallel to the continental slope. The existence of the zones is determined by the contact of different water masses with the bottom. These zones are:
zone with sharp seasonal fluctuations of temperature (warming in summer, cool in winter);
zone with constant low off-bottom temperature (bottom layer of residual winter cooling);
zone warmed up by deep waters of the continental slope.
Every water mass, and, hence, every zone, is characterized by its own shrimp species. Zone (i) occupies the sandy coastal areas of not more than 50 m in depth; Crangon dalli is the most common species of shrimp. Zone (ii) extends from 50 m to 70 to 100 m; the bottom is mainly muddy, and the cold-water forms, Eualus macilentus and Pandalus goniurus, are the most typical species. Thermal conditions favouring P. borealis are observed only in zone (iii) (deeper than 70 to 100 m), and it is the dominant species in this zone. Thus, in the Bering Sea winter cooling is a significant limiting factor in the distribution of P. borealis, which is largely restricted to the zone where cooling is minimal.
Despite the wide distribution of P. borealis in the zone warmed up by deep water, its commercial concentrations occur only off the Pribilof Islands. The formation of the Pribilof population is determined by the combined action of three factors: a favourable thermal regime, since the area is greatly warmed up by deep waters; the presence of a large muddy area; and the closed water circulation (Fig. 6). As a result of this last factor, shrimp larvae do not drift into unfavourable regions for development.
Although the P. borealis is widely distributed in zone (iii), its commercial concentrations are not observed in the warmest layers of the zone, where it might be expected. The shrimps tend to concentrate in the upper layers of the zone, close to the zone of constant low off-bottom temperature. Hence, the Bering Sea shrimp concentrations are found in the zone of interaction of two water masses.
The position of borders between the zones (ii) and (iii) is variable. Due to the winter cooling, zone (ii) enlarges and the zone (iii) decreases correspondingly. Consequently, in winter the Pribilof population migrates 30 to 40 miles southwestward, approaching the continental slope, moving from the summer depths (85 to 100 m) to a depth of 95 to 120 m. In severe winters, cold waters with the temperature below zero can even cover areas of shrimp concentrations. Since a very flat bottom is characteristic of the Pribilof area, shrimp distribution depends very little on the bottom relief, and the influence of hydrological conditions on their distribution becomes especially apparent.
The Pribilof population differs from others in a number of respects. It is the only concentration located at a long distance from the continent; it occupies a very large area; heavy concentration of shrimps can be recorded there; shrimp distribution is almost independent of the bottom relief, and the shrimps carry out rather large seasonal migrations of 30 to 40 miles (56 to 74 km).
3.2 The Gulf of Alaska
In contrast to the Bering Sea, winter cooling is not an important limiting factor for the P. borealis distribution in the Gulf of Alaska. Complex relief and bottom deposits are of much greater importance than in the Bering Sea. Relief and deposits greatly disguise the dependence of shrimp distribution upon water masses, but this dependence is distinctly manifested in the comparatively well investigated Shumagin region.
Comparing shrimp distribution with the off-bottom isotherms, one can note that the maximum catches of shrimps are usually in the zone of sharp temperature gradients, where the mixing of different water masses apparently takes place. Besides, off the Shumagin Islands shrimps avoid the zone where a warm intermediate layer (apparently, a branch of warm Alaska current) is in contact with the bottom.
P. borealis appears to show a general tendency to form concentrations in the zone of interaction of water masses of different origin, and not in waters with some optimum temperature.
The U.S.A. and U.S.S.R. surveys in the Gulf of Alaska (Greenwood, 1959; Johnson, 1959; Ronholt, 1963; Rathjen and Yesaki, 1966; Ivanov, 1963, 1964) supplement each other well, because the American surveys were conducted mainly within the 12 mi (22 km) zone and the Soviet surveys have all been outside this zone. These surveys show that it is in the west part of the Gulf (off the Kodiak and, particularly, Shumagin Islands) that the largest stocks of shrimps occur, both in the inshore and offshore waters.
Apparently, the comparatively wide shelf of the western Gulf, with numerous muddy grooves and hollows, is more favourable for the formation of concentrations than the narrow rocky fjords of the southeastern part of the Gulf. Both islands and bays help to create the circulation system which prevents larvae from drifitng away into unfavourable areas. This makes the western part of the Gulf more favourable for P. borealis than the central part and it is the most commercially important area.
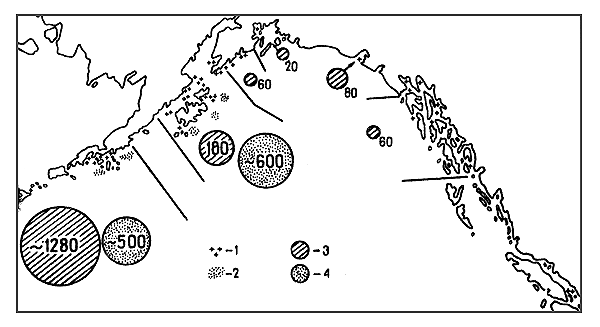
Fig. 7 Inshore and offshore shrimp fishing areas in the Gulf of Alaska and catches from these areas; data on inshore grounds from Ronholt (1963) and Butler (1964), data on offshore grounds original:
| 1 areas of inshore concentrations, | 2 areas of offshore concentrations, |
| 3 average catch kg/h of inshore areas, | 4 average catch kg/h of offshore areas. |
The northern shrimp commercial concentrations in the Atlantic and Pacific can be classified into three types in relation to their remoteness from the continent:
fjord and bay concentrations in the close vicinity of the coast;
offshore concentrations within a short distance, not farther than 30 to 40 mi (56 to 74 km) of the coastline;
offshore concentrations at long distance from the coast.
The first type comprises concentrations in fjords of the Scandinavian countries and those in the coastal zone of the Gulf of Alaska. The second one involves the concentrations off the East and West Shumagin, South and North Kodiak in the Pacific and those off the Skagerrack. The third includes only the Pribilof concentrations in the Bering Sea.
Allen, J.A., 1959 On the biology of Pandalus borealis Kr., with reference to a population off the Northumberland Coast. J.mar.biol.Ass.U.K., 38(1):189–220
Arsenyev, V.S. and Shcherbinin, A.D., 1963 Investigation of currents in the Aleutian waters and in the Bering Sea. Results of researches on the program of the International Geophysical Year. Okeanol.Issled., (8):58–66
Butler, T.H., 1964 Growth, reproduction and distribution of pandalid shrimps in British Columbia. J.Fish.Res.Bd Can., 21(6):1403–52
Greenwood, M., 1959 Shrimp exploration in Central Alaskan waters by m/v John N. Cobb, July-August 1958. Comml.Fish.Rev. 20(7):1–19
Horsted, S. and E. Smidt, 1956 The deep-sea prawn (Pandalus borealis kr.) in Greenland waters. Meddr Danm.Fisk.-og Havunders., N.S., 1(11):1–118
Ivanov, B.G., 1963 Some data on the life history of the shrimp in the western part of the Gulf of Alaska. Trudy vses.nauchno-issled.Inst.morsk.ryb.Khoz.Okeanogr., 48:207–18 (in Russian)
Ivanov, B.G., 1964 On the biology and distribution of shrimp in the Gulf of Alaska and the Bering Sea in winter. Trudy vses.nauchno-issled.Inst.morsk.ryb.Khoz.Okeanogr. 53:185–98 (in Russian)
Johnson, H.C., 1959 King crab, shrimp and bottom fish explorations conducted in certain waters from Shumagins Islands to Unalaska, Alaska, by M/V TORDENSKTOLD, summer and fall, 1957. Comml.Fish.Rev., 21(3):7–19
Kryndin, A.N., 1964 The seasonal and yearly changes in ice conditions and the positions of ice edge in the Far East Seas in relation to the peculiarities in the atmospheric circulation. Trudy gas. okeanogr.Inst., 71 (in Russian)
Rasmussen, B., 1953 On the geographical variation in growth and sexual development of the deep-sea prawn (Pandalus borealis Kr.). FiskDir.Skr.(Havundersok.), 10(3):160 p.
Rathjen, W.F. and M. Yesaki, 1966 Alaska shrimp explorations, 1962–64. Comml.Fish.Rev., 28(4):1–14
Roncholt, L.L., 1963 Distribution and relative abundance of commercially important pandalid shrimps in the northeastern Pacific Ocean. Spec.scient.Rep.U.S.Fish Wildl Serv.(Fish.), (449): 28p.
Anon., 1965 Shrimp life history studies. Comml.Fish.Rev., 27(11):17–8
![]()
![]()
![]()