![]()
![]()
![]()
Les populations de corégones qui se maintiennent de manière autonome nécessitent des eaux oligotrophes ou mésotrophes, fraîches et bien oxygénées. C'est la raison pour laquelle leur aire de répartition en Europe s'étend de l'Europe centrale du nord jusqu'au nord de la Scandinavie et des lles Britanniques à l'ouest à la limite orientale de l'Europe en Russie. En Europe centrale, ils sont abondants dans les lacs subalpins de France, d'Allemagne, d'ltalie, de Suisse, d'Autriche et dans les réservoirs de la République tchèque et slovaque.
A l'extrémité sud de leur aire de répartition, les lacs qu'ils préfèrent sont profonds mais, au nord de l'Europe, les corégones vivent également dans les lacs peu profonds. Pour le maintien autonome des stocks, la température et l'oxygène sont les paramètres de qualité d'eau les plus importants; il en va de même de la présence de substrats appropriés au dépôt du frai. La température maximale létale pour le lavaret est d'environ 22°C, l'optimum étant < 15°C et, pour le peled, ces taux sont un peu plus élevés. Le taux d'oxygène ne devrait pas descendre en dessous de 4 mg/l et la saturation en oxygène sur les frayères devrait être supérieure à 70%. Le degré de sensibilité à une qualité défavorable de l'eau dépend du stade de développement et de la condition physiologique du poisson, du temps d'acclimatation, de la composition chimique de l'eau et encore d'autres facteurs.
La qualité de l'eau n'est pas le seul facteur abiotique qui, en conditions naturelles, affecte l'effectif de la classe d'âge de l'année. Les conditions climatiques (la température, le vent, les rayons du soleil, etc.) sont parmi les facteurs les plus importants pouvant affecter le recrutement des corégones. La sensibilité du poisson au stress environnemental décroît généralement en fonction de sa taille (stade de développement). On peut également repeupler des lacs et des réservoirs où les corégones ne peuvent pas maintenir une population naturelle autonome; c'est le cas dans les lacs eutrophes.
La compétition au sein des cohortes et entre les cohortes est importante et l'on a observé occasionnellement du cannibalisme.
Une relation entre géniteurs et recrues n'a pu être observée que rarement pour la petite marêne chez qui la variabilité de recrutement est grande. En eau oligotrophe, la compétition au sein d'une cohorte au stade larvaire (croissance et survie dépendantes de la densité) est, chez la petite marêne, le mécanisme réputé comme affectant le plus l'effectif de la classe d'âge. La compétition asymétrique entre les petites marênes jeunes et adultes est également considérée comme significative. Dans les lacs eutrophes, les facteurs environnementaux (indépendants de la densité) sont plus importants.
On a rarement pu mettre en relation le recrutement dans les stocks naturels de lavaret et la taille de la population. Pour cette raison, on considère que, dans la plupart des cas, le recrutement des lavarets est déterminé par des mécanismes environnementaux abiotiques et biotiques indépendants de la densité. Ceci explique le succès des repeuplements finlandais en juvéniles dans des lacs contenant des populations de lavarets indigènes. Les lavarets sont connus pour leur prédation considérable sur leurs propres oeufs et larves, ce qui pourrait jouer un rôle dans la régulation de la population.
II faut se rappeler que lors de repeuplements en larves vésiculées, les mêmes facteurs environnementaux agissent autant que pour les larves écloses naturellement. Ceci explique aussi, au moins partiellement, pourquoi les repeuplements à partir de larves vésiculées échouent si fréquemment en présence de stocks à reproduction naturelle dans le lac. Contrairement aux repeuplements avec des juvéniles, il n'est donc pas réaliste d'espérer quelque résultat positif marqué de repeuplements avec des larves vésiculées de lacs dans lesquels des populations à reproduction naturelle existent déjà. Ceci est valable pour tous les corégones. La situation est différente dans les lacs eutrophes dans lesquels la pêche est efficace et les conditions nutritives bonnes.
II est logique de supposer que des mécanismes compensatoires peuvent également agir dans une certaine mesure lors de repeuplements en poissons juvéniles. Dans les systèmes naturels, le nombre de recrues est généralement déterminé avant la période juvénile et donc les repeuplements en juvéniles accroissent la densité de la population et donc aussi la compétition intraspécifique pour les ressources. Dans de telles situations, le nombre de recrues est déterminé par des mécanismes dépendants (compensatoires) de la densité. La croissance des juvéniles dépend en particulier de la taille de la cohorte. Un retard de la croissance accroît vraisemblablement la vulnérabilité à la prédation et affecte ainsi le nombre de recrues pour la pêche.
Pour cette raison, on pense que dans les lacs peu eutrophes qui possèdent des stocks de lavaret se reproduisant naturellement, les repeuplements avec des poissons juvéniles tendent à être plus efficaces que ceux effectués avec des larves. Ceci est probablement valable également pour le peled, bien que cela ne figure dans les observations reprises ultérieurement (au tableau 1). II faut y noter la grande amplitude des valeurs présentées et admettre que ces valeurs devraient utilement être publiées pour les recherches à venir. Dans les lacs plus eutrophes dépourvus de population naturelle de corégones, la taille des sujets de repeuplement est moins importante.
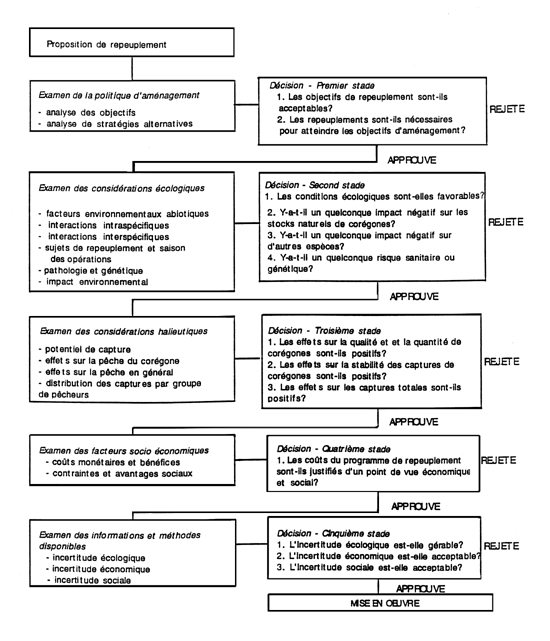
Figure 1. Un modèle pour la planification des repeuplements en corégones. Les examens, à gauche, illustrent les différents niveaux de la collecte des données et les champs de décision, à droite, les stades de décision respectifs, avec des questions qui s'y rapportent. Les repeuplements devraient être rejetés si les réponses sont négatives.
Les interactions entre les espèces de poissons sont très complexes et peuvent changer aux différents stades de leur vie. Les interactions peuvent se présenter sous la forme d'une compétition pour les ressources ou sous la forme d'une prédation.
La petite marêne est nettement planctonophage et supplante le peled et le lavaret dans les lacs oligotrophes. II a été démontré en Scandinavie que, dans les lacs abritant le lavaret, des repeuplements avec des petites marênes ont de bonnes chances de réussite et ce, quel que soit leur stade de développement; pour des repeuplements avec des larves vésiculées de lavaret, le contraire est rarement exact. Cependant, lorsque des lavarets à un stade plus avancé sont utilisés, les succès sont plus fréquents.
Le peled semble sensible à la compétition alimentaire dans les lacs oligotrophes où la petite marêne et le lavaret sont tous deux présents. Les captures provenant de repeuplement en peled n'ont dans ce cas jamais été bonnes et ce, quelle que soit la taille ou l'âge des peleds déversés. La tendance générale pour les espèces qui possèdent un nombre plus élevé de branchiospines est d'occuper la zone pélagique dans laquelle elles se nourrissent, alors que les espèces qui possèdent un nombre plus faible de branchiospines restent en zone littorale et s'y nourrissent de benthos. Ce problème n'a pas été rencontré dans les lacs eutrophes.
L'introduction et le repeuplement en lavaret a été dommageable pour les populations d'ombles chevaliers dans de nombreux lacs scandinaves. La pérennité de l'omble chevalier et du lavaret est cependant possible dans les grands lacs oligotrophes. Les larves de lavaret sont la proie des perches et, dans de nombreux petits lacs, le lavaret a été exterminé par la perche. Cependant, on peut limiter l'action des perches dans les lacs de faible étendue par l'introduction de lavarets à un stade de développement plus avancé. Néanmoins, les grands lacs plus profonds procurent un avantage au lavaret par rapport à la perche. La tendance générale est que le lavaret domine la perche dans les grands lacs profonds comportant une zone pélagique étendue et que la perche tend à dominer la lavaret dans les plus petits lacs, plus chauds. Les mécanismes de dominance sont valables également pour les stades jeunes de développement.
Les espèces prédatrices (brochet, lote de rivière, truite fario, etc.) se nourrissent de corégones. La prédation dépend de la taille et, en conséquence, la mortalité naturelle est très importante chez les individus de plus petite taille. Le risque de prédation décroît en fonction de la taille du poisson. Plus les repeuplements sont importants, plus les conditions sont favorables aux prédateurs. La détermination du niveau de prédation sur les oeufs et les larves peut d'ailleurs servir à estimer le taux de recrutement des corégones dans de nombreux lacs.
Certains effets positifs ont été observés dans des lacs polonais légèrement eutrophes dans lesquels les lavarets issus des repeuplements ont tendance à remplacer les petits cyprinidés dans les communautés piscicoles et améliorent de cette façon la qualité de l'environnement et la valeur des captures. L'impact peut alors s'avérer similaire à celui résultant de l'élimination massive d'espèces indésirables. Certaines observations indiquent que pour des lacs dans lesquels les stocks de petite marêne et de lavaret sont constitués de spécimens de grande taille et où l'abondance de la population est restée très faible pendant plusieurs années, des éliminations massives d'espèces indésirables peuvent induire une augmentation très importante des populations de petite marêne et de lavaret.
Aux endroits où des repeuplements d'hybrides de peled et de lavaret ont été effectués dans des lacs eutrophes, soit à partir d'écloseries, soit par des hybridations naturelles, la croissance de la population s'est avérée faible, la reproduction précoce et la valeur des captures faible.
Il est très important de prendre en compte les facteurs écologiques pour déterminer les sujets à déverser (espèce, stade de développement, taille, etc.) et la saison des opérations de repeuplement. La sensibilité aux manipulations dépend des conditions physiologiques, de la taille du poisson destiné au repeuplement et de la température ambiante. Il est évident que plus le poisson destiné au repeuplement est gras, moins il est résistant et que plus la température est élevée, plus il est sensible aux manipulations. La taille des poissons de repeuplement doit être déterminée sur base des conditions environnementales (exigences physiologiques quant à la qualité de l'eau, composition des espèces, prédation, etc.).
L'environnement physique et le taux de prédation varient en fonction des saisons et doivent être pris en considération pour déterminer la période appropriée pour les opérations de repeuplement. D'autre part, la période des opérations de repeuplement doit être déterminée sur base des cycles de production et des périodes de repeuplement techniquement souhaitables. Les transferts pour les petites marênes ne devraient s'effectuer qu'en hiver quand les jeunes de l'année sont faciles à récolter et que le transport peut être organisé avec un minimum de pertes de poisson.
Les corégones peuvent être porteurs de nombreuses maladies bactériennes, virales ou parasitaires propres aux salmonidés. La probabilité de ce portage s'accroît normalement avec l'âge et la taille. Les parasites les plus observés sont Diplostomum, Proteocephalus et Triaenophorus dans les élevages en étang et en écloserie. Les populations denses peuvent accroître la sensibilité à certains parasites qui sont susceptibles de déprécier la valeur des captures. Triaenophorus est particulièrement problématique là où les brochets sont abondants; la valeur marchande des poissons parasités peut alors être pratiquement nulle. Pour cette raison, il est très important d'utiliser dans la mesure du possible du matériel de repeuplement porteurs de charges parasitaires minimales.
Lorsque les poissons provenant d'écloserie sont déversés, ils peuvent avoir un effet génétique néfaste à cause de la consanguinité, l'hybridation et l'introgression dans les populations naturelles de corégones. Il peut en découler une perte de variabilité génétique (caractère hétérozygote réduit) et un accroissement des gènes “d'écloserie” non adaptés et, par conséquent, une réduction de la tenue et la vigueur générale des stocks de corégones, mais très peu d'informations sont disponibles sur le sujet.
Les risques génétiques sont peu importants dans les lacs où les stocks de corégones ne se reproduisent pas naturellement mais ils peuvent être très importants dans les lacs oligotrophes où les stocks sont naturels. Le matériel local doit toujours être utilisé là où c'est possible. Le transfert de corégones vers un nouvel environnement ou vers de nouvelles aires géographiques dans lesquelles des populations de corégones indigènes existent déjà est risqué, s'il est destiné à produire une population qui se maintient d'elle-même dans la mesure où la relation entre la période de reproduction et les conditions environnementales semble être, au moins partiellement, déterminées génétiquement; les hybrides pourraient ainsi être moins bien adaptés que les stocks d'origine.
Les risques génétiques et pathologiques qui entrent en jeu dans les repeuplements peuvent être réduits en respectant les consignes suivantes:
Afin de maintenir une base solide pour le matériel de repeuplement, les lacs qui possèdent une souche locale utilisée pour la récolte de frai destiné à l'élevage en écloserie ne devraient pas être repeuplés avec d'autres matériels que la souche locale.
Les lavarets provenant de régions marines ne devraient pas être introduits dans les eaux continentales et les oeufs de lavarets marins devraient être désinfectés soigneusement lorsque l'incubation se déroule en eaux douces.
Le transport de poissons d'un réseau hydrographique à l'autre devrait être réduit au strict minimum.
L'origine des poissons utilisés pour le repeuplement, leur condition et les risques sanitaires éventuels doivent être examinés avant le repeuplement.
Pour éviter une reproduction sélective non intentionnelle, il est important d'utiliser tout l'éventail de la population reproductrice. Par exemple, la récolte de trop de matériel tôt dans la saison de reproduction favorise la maturité précoce et par conséquent les plus petits poissons.
Lors de repeuplements intensifs de petite marêne et de peled, on peut s'attendre à un accroissement de la prolifération des algues et de la biomasse, de par leur forte prédation sur le zooplancton; pour cette raison, leur utilisation n'est pas toujours considérée comme favorable dans les réservoirs d'eau potable. Cependant, il est évident qu'un repeuplement intensif en petite marêne peut améliorer la qualité de l'environnement et ralentir le processus d'eutrophisation. Ceci est dû au fait que les captures de petite marêne provenant de repeuplements peuvent être très importantes et que des quantités substantielles de phosphore sont retirées de l'écosystème à l'occasion de la capture de poissons. Pour cette raison, le repeuplement en petite marêne a également un effet bénéfique indirect sur le stock de poissons dans les lacs. Il existe des lacs dans lesquels l'équilibre écologique est maintenu principalement grâce aux repeuplements importants en petite marêne et grâce à une pression de pêche importante (par exemple, le Lac Maroz). La contradiction apparente de l'expérience polonaise face aux résultats des biomanipulations s'explique par le fait que la pêche dans les lacs polonais est une pêche très efficace portant sur plusieurs espèces (en fait une expérience de biomanipulation à grande échelle).
![]()
![]()
![]()