![]()
![]()
![]()
Un certain nombre de facteurs ou de combinaisons de facteurs influent sur la santé des poissons au cours du transport et sur leur survie.
La qualité est un élément déterminant: les poissons doivent être sains et en bon état. Les individus affaiblis devraient être éliminés du lot delivraison notamment lorsque la température est élevée pendant le transport. Dès l'instant où les poissons sont de mauvaise qualité, en réduire fortement la densité dans les conteneurs ne suffit pas pour éviter des pertes. Le taux de mortalité des individus débilités est nettement plus élevé que celui des poissons sains, lorsque la durée du transport est allongée.
Il peut également être nécessaire d'habituer préalablement les poissons à une eau dont la température est plus basse. Pour refroidir l'eau, Il vaut mieux utiliser de la glace naturelle que de la neige carbonique. A titre indicatif on considère qu'avec 25 kg de glace on peut abaisser de 2°C la température de 1 000 litres d'eau. Si l'eau que l'on refroidit contient déjà des poissons, il faut veiller à ce que sa température ne tombe pas de plus de 5°C à l'heure, et à ce que les poissons ne soient pas au contact direct de la glace. L'écart total de température ne devrait pas excéder 12–15°C, compte tenu de l'espèce et de l'âge des individus (recommandation RFA, 1979).
Sauf au stade larvaire, tous les poissons devraient, avant le transport, rester à jeun au moins pendant un jour. Dans des conditions identiques, la durée possible du transport est réduite de moitié lorsque le tube digestif n'est pas totalement vide (Pecha, Berka and Kouril, 1983; Orlov et al., 1974) car la consommation d'oxygène est accrue tout comme la sensibilité au stress. En outre les excréments produits absorbent la plupart de l'oxygène de l'eau. Toutefois, dans le cas des larves, il convient de tenir compte de leur résistance au jeûne. Ainsi les larves des poissons herbivores ne peuvent pas rester sans nourriture plus de 20 heures et l'endurance de nombreuses espèces d'aquarium est inférieure à 12 heures (Orlov, 1971).
Bien qu'il soit fondamental d'assurer une teneur suffisante en oxygène dissous au cours du transport, la présence d'une forte quantité de cet élément dans un bac ne garantit pas forcément que les poissons sont en bon état. En effet, leur faculté d'utiliser l'oxygène dépend de leur résistance au stress, de la température de l'eau, du pH et des concentrations de gaz carbonique et de produits du métabolisme comme l'ammoniac.
Le poids des poissons et la température de l'eau sont les facteurs déterminants qui influent sur le métabolisme et sur la consommation d'oxygène pendant le transport. Les poissons ont d'autant plus besoin d'oxygène qu'ils sont plus gros et que l'eau est plus chaude. Si la température de l'eau augmente de 10°C (de 10° à 20°C par exemple) la consommation d'oxygène est pratiquement doublée. Pour chaque augmentation de 0,5°C, il faudrait diminuer d'environ 5,6 pour cent le chargement et, inversement, pour toute baisse de 0,5°C, on peut accroître le chargement dans les même proportions (Piper et al., 1982).
L'agitation des poissons après avoir été manipulés accroît aussi la consommation d'oxygène. La demande d'oxygène est en l'occurence de 3 à 5 fois plus élevée et il faut par exemple aux alevins de salmonidés plusieurs heures pour retrouver leur niveau normal de métabolisme de l'oxygène, c'est-à-dire en général après être arrivés à destination (Lusk et Krcál, 1974).
Placé dans de l'eau contenant de l'oxygène en quantité illimitée un poisson au repos n'en consomme d'ordinaire qu'un minimum. Pendant le transport, il en consomme davantage car il n'est plus au repos. En outre, s'il est agité au moment du chargement ou dérangé en cours de transport, il se peut qu'il consomme près de la quantité maximum.
La quantité d'oxygène utilisée dépend aussi de la quantité disponible. Lorsque la teneur est élevée, la consommation s'établit à un niveau constant. Lorsqu'elle est faible, le poisson en utilise moins, quel que soit son niveau d'activité.
Dans les systèmes de transport l'eau a souvent une teneur en oxygène qui ne permet pas de couvrir les exigences des poissons. Pour compenser ce manque, le poisson modifie son métabolisme de manière à utiliser l'oxygène accumulé dans son organisme. C'est un peu le cas d'un homme qui est au repos et qui accomplit soudain un gros effort avant d'avoir emmagasiné la quantité d'oxygène nécessaire. Il se créé donc pour l'homme comme pour le poisson une dette d'oxygène qui devra être soldée lorsque les conditions favorables se présenteront.
Les poissons ont particulièrement besoin d'oxygène pendant l'heure qui suit le chargement. Comme ils sont agités il leur en faut de grandes quantités et ils ont peu de temps pour s'adapter. La demande d'oxygène varie aussi beaucoup d'une famille de poissons à l'autre. Ainsi, comme le relève Uryn (1971), lorsque la température de l'eau passe de 4° à 14°C au cours du transport, les alevins de Coregonus lavaretus consomment 2,4 fois plus d'oxygène que ceux de C. albula. La taille du poisson est également importante. Un gros poisson consomme moins d'oxygène par unité de poids qu'un petit. Pour la plupart des poissons d'eau chaude, la teneur en oxygène de l'eau devrait être supérieure à 5 mg 1-1 en conditions normales afin d'éviter que le manque d'oxygène ne devienne un facteur de stress important.
La recommandation de la RFA (1979) mentionne certains coefficients de conversion pour la demande d'oxygène. Ainsi 25 kg de truites arc-en-ciel pesant chacune 250 g ont la même demande d'oxygène que 20 kg de truites de peuplement (1 100 poissons), que 17 kg de truites de peuplement de 8 cm (3 200 poissons) ou que 12 kg d'alevins forcés d'une longueur de 4 cm (environ 23 000 poissons). Si l'on pose que la demande d'oxygène de la carpe est égale à 1, les niveaux de demande d'oxygène des autres poissons après conversion sont les suivants:
| truite | 2,83 | brème | 1,41 |
| sandre | 1,76 | brochet | 1,10 |
| gardon | 1,51 | anguille | 0,83 |
| perche | 1,46 | tanche | 0,83 |
D'après Schevchenko (1978) la consommation d'oxygène de Coregonus peled par kilo et par heure à la température de 10°C est de 100 mg; cette valeur est de 68 mg pour l'esturgeon, de 50 à 60 mg pour le brochet et de 45 mg pour les carpes de 500–600 g.
Avec le transport en sac fermé, la teneur en oxygène de l'eau n'est pas en général un facteur limitatif, car le sac en contient assez sous pression. Exceptionnellement, l'oxygène peut venir à manquer si par exemple la densité des poissons est trop élevée ou si le transport dure plus longtemps qu'ils ne peuvent le supporter. Les poissons morts soustraient de l'oxygène aux vivants et favorisent la multiplication des bactéries qui en consomment beaucoup, multiplication qui, à son tour, risque d'engendrer des métabolites toxiques. Le mucus produit par les poissons fournit un autre substrat au développement bactérien qui fait baisser encore la teneur en oxygène de l'eau. Ce processus s'accentue avec l'élévation de la température de l'eau.
Une eau à forte teneur en oxygène n'a pas d'effet négatif sur les poissons. Ainsi la limite pour la truite arc-en-ciel est de 35 mg par litre, teneur qu'il est impossible d'obtenir en pratique comme le mentionne Heiner (1983). Les poissons sont en général capables de régler le volume d'oxygène qui pénètre dans leur organisme, mais il existe peut-être des exceptions. Pour le moment, on ne sait pas quels effets pourrait avoir sur les larves une exposition prolongée à une eau à forte teneur en oxygène et à température élevée. A ce stade en effet les poissons n'ont pas la faculté de maintenir à un niveau optimal l'oxygène de leur sang.
Dans les systèmes fermés, de légères vibrations du sac permettent à l'oxygène atmosphérique de pénétrer dans l'eau. Au cours d'arrêts prolongés, lorsque les sacs restent immobiles, il peut arriver que des poissons meurent bien que le niveau de la réserve d'oxygène soit encore satisfaisant. Ainsi en va-t-il surtout avec des stocks denses de salmonidés qui ont besoin de beaucoup d'oxygène, mais non avec des cypriniés (à l'exception des alevins) parce que les poissons de cette espèce agitent eux-mêmes l'eau dans les sacs et établissent ainsi un contact suffisant avec l'oxygène de l'atmosphère. Le tableau 1 indique combien de temps les alevins de salmonidés peuvent rester dans des sacs fermés immobiles avant que les concentrations d'oxygène n'arrivent à un seuil critique.
Tableau 1
Nombre d'heures de transport que peuvent supporter les alevins de salmonidés dans des sacs fermés immobiles avant que les concentrations d'oxygène n'arrivent à un seuil critique
| Poids individuel moyen (g) | Température (°C) | Poids total des poissons (kg) | ||||||
| 0,25 | 0,5 | 1,0 | 1,5 | 2,0 | 2,5 | 3,0 | ||
| 0,5 | 5 | 10,0 | - | - | - | - | - | - |
| 10 | 10,0 | - | - | - | - | - | - | |
| 15 | 6,0 | - | - | - | - | - | - | |
| 20 | 3,9 | - | - | - | - | - | - | |
| 1–2 | 5 | 10,0 | 10,0 | - | - | - | - | - |
| 10 | 10,0 | 6,0 | - | - | - | - | - | |
| 15 | 6,7 | 3,3 | - | - | - | - | - | |
| 20 | 4,4 | 2,2 | - | - | - | - | - | |
| 5–10 | 5 | 10,0 | 10,0 | 7,3 | 4,7 | - | - | - |
| 10 | 10,0 | 6,6 | 3,2 | 2,1 | - | - | - | |
| 15 | 8,0 | 3,9 | 1,9 | 1,2 | - | - | - | |
| 20 | 5,3 | 2,6 | 1,3 | 0,8 | - | - | - | |
| 20–50 | 5 | 10,0 | 10,0 | 9,1 | 5,9 | 4,3 | - | - |
| 10 | 10,0 | 7,3 | 3,5 | 2,3 | 1,7 | - | - | |
| 15 | 9,2 | 4,5 | 2,2 | 1,4 | 1,0 | - | - | |
| 20 | 5,8 | 2,8 | 1,4 | 0,9 | 0,7 | - | - | |
| 100 | 5 | 10,0 | 10,0 | 10,0 | 6,7 | 4,9 | 3,8 | 3,1 |
| 10 | 10,0 | 8,2 | 4,0 | 2,6 | 1,9 | 1,5 | 1,2 | |
| 15 | 10,0 | 4,9 | 2,4 | 1,5 | 1,1 | 0,9 | 0,7 | |
| 20 | 6,5 | 3,2 | 1,5 | 1,0 | 0,7 | 0,6 | 0,5 | |
La qualité de l'eau détermine la charge de poissons et la durée du transport. L'eau utilisée doit être testée avant l'expédition d'un lot important. Le pH de l'eau est un élément de contrôle parce que la teneur en ammoniac et en CO2 en dépendent directement (figure 1).
Avec l'allongement de la durée du transport, le CO2 produit par la respiration des poissons acidifie le pH de l'eau (un pH de 7–8 est considéré comme optimal). Les modifications rapides du pH stressent les poissons mais on peut utiliser des solutions tampons pour les stabiliser pendant le transport. Le trishydroxylméthyl aminométhane est une solution tampon organique très efficace aussi bien en eau douce qu'en eau de mer. Elle est très soluble, stable et s'emploie facilement. Cette solution a été utilisée pour 29 espèces de poissons et n'a eu aucun effet nocif. Il est recommandé de l'appliquer à des doses de 1,3 à 2,6 g par litre pour les transports courants (Piper et al., 1982).
De fortes concentration de gaz carbonique sont nocives pour les poissons et peuvent en entraver le transport. Le CO2, dégagé par la respiration des poissons et des bactéries, acidifie l'eau et bien qu'il réduise la teneur en ammoniac non-ionisé il diminue aussi la capacité du sang en oxygène. Un niveau élevé de CO2 peut entraîner la mort des poissons même si celui de l'oxygène est apparemment suffisant. Il semble que la truite tolère un niveau de gaz carbonique inférieur à 15 mg 1-1 si l'alimentation en oxygène est satisfaisante et la température raisonnable, mais donne des signes d'épuisement lorsque la teneur en gaz carbonique avoisine 25 mg 1-1 (Piper et al., 1982).
Les poissons transportés dans des bacs sont soumis à des niveaux toujours plus élevés de gaz carbonique, qui peuvent dépasser 20–30 mg. 1-1, si l'aération laisse à désirer. En général pour chaque millilitre d'oxygène qu'il absorbe un poisson produit environ 0,9 ml de CO2. Si le niveau de CO2 augmente rapidement, lorsque les poissons sont très nombreux par exemple, on note des signes de détresse. De fortes concentrations de CO2 sont cependant tolérées si l'accumulation se fait lentement.
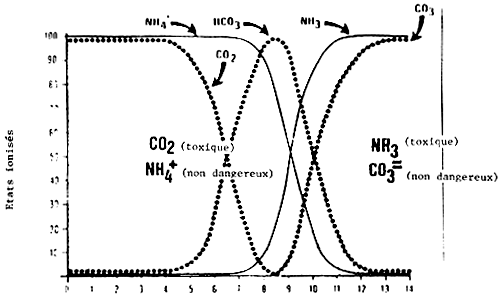
Figure 1 Niveau d'ammoniac et de gaz carbonique (en pourcentage) pour les divers niveaux de pH (Amend et al., 1982)
Il est indispensable d'assurer une bonne ventilation des unités de transport. Dans des conteneurs bermétiquement fermés sur le dessus une forte accumulation de CO2 peut se produire qui stresse les poissons. L'aération de l'eau réduit les concentrations de CO2 dissous pourvu que la ventilation soit bonne.
Comme l'indiquent Pecha, Berka et Kouril (1983), en espaces clos les concentrations critiques de CO2 varient de 140 ml par litre pour les poissons thermophiles à près de 40 ml par litre pour ceux qui préfèrent l'eau froide. Kruzhalina, Averina et Vol'nova (1970) donnent des niveaux critiques de CO2 plus précis pour les systèmes fermés à savoir: 60–70 ml par litre pour les salmonidés; 40 ml par litre pour les esturgeons adultes, 20 ml par litre pour les alevins d'esturgeon, 140–160 ml pour les poissons herbivores adultes, 100 ml par litre pour les alevins de poissons herbivores et 80 ml par litre pour les larves des espèces herbivores. Toutes ces données concernent les systèmes fermés; dans les systèmes ouverts, le CO2 est expulsé de l'eau par n'importe quel dispositif d'aération. En conteneur fermé le seuil critique de CO2 a beaucoup moins d'importance dès l'instant où le nombre des poissons diminue.
La concentration de chlore dans l'eau est un autre facteur important, bien que - comme le gaz carbonique - le chlore puisse être éliminé par l'aération de l'eau. On considère qu'une concentration de 0,5 mg par litre est dangereuse, mais des niveaux même inférieurs, par exemple 0,2 mg par litre, gênent considérablement le système respiratoire du poisson (Shevchenko, 1978).
Pendant le transport, l'ammoniac (NH3) s'accumule dans l'eau sous l'effet du métabolisme protéinique des poissons et de l'action des bactéries sur les déchets. Si l'on réduit le métabolisme des poissons en abaissant la température de l'eau, on diminue l'activité, et par conséquent la production de NH3. Pour que les bactéries en dégagent moins, on peut ne transporter que des poissons à jeun, dont l'estomac et l'intestin sont vides.
La température et l'heure de la dernière prise de nourriture jouent un grand rôle dans la régulation de l'excrétion d'ammoniac. Ainsi dans une eau à 1°C les truites rejettent 66% d'ammoniac en moins que dans une eau à 11°C et les individus qui ont jeûné pendant 63 heures avant l'expédition en dégagent moitié moins que les poissons récemment nourris. Les poissons dépassant 10 cm devraient jeûner au moins 48 heures, ceux de 20 cm et plus 72 heures (Piper et al., 1982).
La quantité d'ammoniac non-ionisé augmente à mesure que s'élèvent la température et le pH de l'eau (tableau 2).
Il n'est pas possible de fournir une valeur maximale admissible car la toxicité de l'ammoniac varie considérablement en fonction de la température de l'eau et du pH. Toutefois, il est rare d'arriver à des concentrations critiques d'ammoniac dans des conditions normales de transport.
Tableau 2
Pourcentage d'ammoniac non ionisé dans l'eau (de 0 à 30°C) pour un pH de 6 à 10 (Emerson et al., 1975)
| Température °C | pH | ||||
| 6,0 | 7,0 | 8,0 | 9,0 | 10,0 | |
| 0 | 0,008 | 0,08 | 0,82 | 7,64 | 45,3 |
| 2 | 0,01 | 0,10 | 0,97 | 8,90 | 49,3 |
| 4 | 0,01 | 0,12 | 1,14 | 10,3 | 53,5 |
| 6 | 0,01 | 0,14 | 1,34 | 11,9 | 57,6 |
| 8 | 0,02 | 0,16 | 1,57 | 13,7 | 61,4 |
| 10 | 0,02 | 0,19 | 1,83 | 15,7 | 65,1 |
| 12 | 0,02 | 0,22 | 2,13 | 17,9 | 68,5 |
| 14 | 0,03 | 0,25 | 2,48 | 20,2 | 71,7 |
| 16 | 0,03 | 0,29 | 2,87 | 22,8 | 74,7 |
| 18 | 0,03 | 0,34 | 3,31 | 25,5 | 77,4 |
| 20 | 0,04 | 0,40 | 3,82 | 28,4 | 79,9 |
| 22 | 0,05 | 0,46 | 4,39 | 31,5 | 82,1 |
| 24 | 0,05 | 0,53 | 5,03 | 34,6 | 84,1 |
| 26 | 0,06 | 0,61 | 5,75 | 37,9 | 85,9 |
| 28 | 0,07 | 0,70 | 6,56 | 41,2 | 87,5 |
| 30 | 0,08 | 0,80 | 7,46 | 44,6 | 89,0 |
La température de l'eau a son importance. Lorsqu'elle est basse, le pH reste élevé et le métabolisme du poisson diminue. En règle générale, on considère les tranches de température suivantes comme optimales pour le transport des poissons: de 6 à 8°C pour les poissons d'eau froide et de 10 à 12°C pour les poissons d'eau chaude, en été; de 3 à 5°C pour les poissons d'eau froide et de 5 à 6°C pour les poissons d'eau chaude au printemps et à l'autonme; et de 1 à 2°C pour tous les poissons en hiver. Evidemment ces tranches de température ne s'appliquent pas aux stades précoces des alevins. Le jeune frai des cyprinidés ne peut pas être transporté à des températures inférieures à 15°C, celui des salmonidés à des températures supérieures à 15–20°C; tandis qu'on considère comme optimale une température de 10°C pour les phases précoces des alevins des corégonidés (Pecha, Berka et Kouril, 1983; Orlov et al., 1971; Shevchenko, 1978).
Il faut aussi tenir compte de l'espace dont disposent les poissons. Pour les alevins, le rapport entre la quantité de poissons transportés et l'eau ne devrait pas dépasser 1:3, pour les spécimens plus gros, comme les reproducteurs, il devrait se situer entre 1:2 et 1:3, mais pour des poissons plus petits, il tombe à 1:100–1:200 (Pecha, Berka et Kouril, 1983).
La recommandation de la République fédérale d'Allemagne indique les rapports suivants entre le poids des poissons et le volume d'eau nécessaire dans un bac (avec une aération satisfaisante, une température de 8–12°C et pour des voyages de courte durée de 1–2 heures): carpe de consommation (1:1) carpe de peuplement (1:1,5), truite arc-en-ciel de consommation (1:3), truite de peuplement (1:4,5), brochet de peuplement (1:2), poissons herbivores (1:2).
Le surmenage et la fatigue des poissons influent également sur les conditions de transport. Lorsque les poissons sont placés dans des conteneurs, ils déploient en général une grande activité musculaire; des muscles sont utilisés intensivement et le sang (donc l'oxygène) vient à manquer. Lorsqu'il y a production d'énergie sans quantité suffisante d'oxygène, un autre mécanisme entre en jeu. L'acide lactique qui s'accumule dans les muscles et dans le sang fait baisser le pH de ce dernier, d'où une moindre consommation d'oxygène. La quantité d'acide lactique accumulée après quelques minutes d'intense activité musculaire reste stable pendant 24 heures. L'excitabilité et le rétablissement à la suite des effets secondaires de l'excitation varient d'une espèce à l'autre. La consommation d'oxygène est plus élevée pendant les 15 premières minutes du transport qu'au cours des périodes successives de 15 minutes (Dupree et Huner, 1984). C'est pourquoi une injection supplémentaire d'oxygène (2 fois supérieure à la normale) devrait être fournie pendant le chargement et la première heure de transport. Le flux d'oxygène peut ensuite être ramené au niveau normal de 6 mg 1-1 après cette période d'acclimatation, une fois que les poissons se sont calmés et que leur consommation d'oxygène s'est stabilisée (Piper et al., 1982).
Le poids total de poissons qu'il est possible de transporter dans un conteneur est d'autant plus réduit que le poids des individus est faible car la consommation d'oxygène est plus forte et le besoin d'espace plus grand (le facteur espace prend nettement plus d'importance) (figure 2).
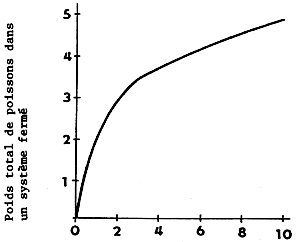
Figure 2 Rapport entre la densité du stock de cyprinidés en système fermé et le poids individuel des poissons lorsque la température de l'eau est de 20°C et la durée du transport de 5 heures (Orlov et al., 1974)
La densité du stock de poissons dans les conteneurs dépend également de la durée du transport (figure 3). Cette relation est représentée par une courbe hyperbolique et non par une ligne droite.
Le rapport entre la densité du stock de poissons dans le conteneur et la température est indiqué à la figure 4. Une température plus élevée nécessite une réduction du poids total du stock.
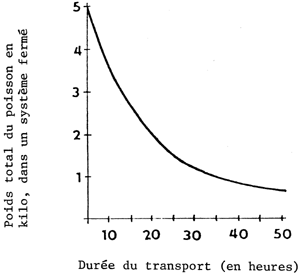
Figure 3 Rapport entre la densité du stock de cyprinides dans un système fermé et la durée du transport: poids individuel: 10 g; durée du transport: 5 heures; température de l'eau: 20°C (orlov et al., 1973)
Pour les cyprinidés la durée du transport est surtout primordiale au stade larvaire. Des trajets de plus de vingt-quatre heures comportent toujours quelques risques, même si par ailleurs toutes les conditions sont bonnes (Pecha, Berka et Kouril, 1983).
La densité des stocks de salmonidés dans un conteneur est toujours inférieure à celle couramment observée pour les cyprinidés car ils consomment plus d'oxygène et résistent moins bien aux concentrations de CO2.
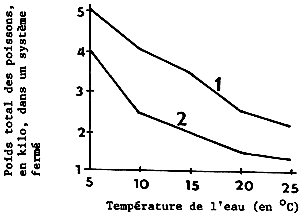
Figure 4 Rapport entre la densité du stock de cypriaidés dans un système fermé et la température de l'eau. 1. poids individuel: 10 g; durée du transport: 15 heures 2. poids individuel: 5 g; durée du transport: 25 heures (Orlov et al, 1974)
Les conditions d'expédition influent aussi sur la composition du sang du poisson et les paramètres biochimiques du sérum sanguin. Une température accrue et un rapport plus faible entre le poids des poissons et la quantité d'eau favorise le développement des globules rouges et une concentration plus élevée d'hémoglobine. Ces modifications ne se produisent pas à des températures plus basses et lorsque la proportion des poissons par volume d'eau est plus faible (Shevchenko, 1978). Carmichael (1984) a également attiré l'attention sur les modifications intervenant pendant le transport dans le sang d'alevins (15–24 cm) de blackbass à grande bouche. Il a constaté qu'avec une densité élevée de tels alevins le niveau des corticoïdes et du glucose dans le plasma montait et se maintenait ainsi même au terme du transport. La mortalité directement liée au transport était faible, mais les effets secondaires imputables au stress entraînaient une mortalité retardée, conséquence d'un disfonctionnement de la pression osmotique, et la maladie.
L'auteur conseille de laisser le blackbass à grande bouche se rétablir au moins pendant 64 heures.
Il convient également de noter que le lâcher des poissons une fois arrivés à destination constitue parfois la phase la plus critique du transport.
Dans l'unité de transport les poissons sont quelque peu stressés et l'immersion soudaine dans une eau dont les caractéristiques sont différentes ou dont la qualité est médiocre aggrave encore leurs troubles, parfois au-delà du seuil de tolérabilité. Par eau de mauvaise qualité, on entend par exemple une eau souterraine pompée peu de temps auparavant et contenant peu d'oxygène ou beaucoup de gaz carbonique. Les différences entre l'eau de l'unité de transport et l'eau d'accueil peuvent porter sur le pH, la température ou la saturation en gaz.
On peut, pour conclure, dégager sur le plan technique et organisationnel, plusieurs observations de la littérature spécialisée. Ainsi, la majorité des auteurs recommande, quelle que soit la densité des stocks conseillée pour le transport, de prendre chaque fois en considération les conditions spécifiques et d'apporter le cas échéant les modifications qui s'imposent. Il est également recommandé d'appliquer une densité qui permette de multiplier la durée du transport au moins par 1,5 afin d'éviter les désagréments pouvant découler d'un contretemps (panne de camion, impossibilité de s'en tenir aux horaires de train ou d'avion). Si les poissons sont transportés pour être acclimatés ou s'il s'agit d'espèces en voie de disparition, la densité du stock devrait être encore réduite. En pareil cas en effet, les considérations économiques passent au second plan; ce qu'il faut c'est garantir un taux de survie de 100 pour cent. Comme cependant on ne saurait jamais faire totalement abstraction de l'aspect économique du transport, lorsque celui-ci coûte cher et que le poisson n'a qu'une assez faible valeur, la densité des poissons peut être accrue car des pertes plus importantes sont admises.
![]()
![]()
![]()