![]()
![]()
![]()
by
R.J. SLACK-SMITH
Department of Fisheries and Fauna
Perth, Western Australia
Abstract
The fishable life span of Penaeids is normally not more than about one year and so the catch for a particular year depends upon the recruits entering the fishery just prior to or during that year. The basic requirement for management is the knowledge of variations in the timing and strength of this recruitment.
Fishery models examined are primarily based on fin fisheries of the North Atlantic and are inadequate for Western Australian penaeid fisheries. The basic assumptions on recruitment, age estimation, growth pattern and effect of migration are inapplicable.
A model is proposed based on the use of detailed data on catch, effort and size composition of catches. This model distinguishes between the effects of growth, migration and variation in density with time. It provides a means of examining time, place and strength of recruit groups entering the fishery.
UN MODELE DESCRIPTIF ET ANALYTIQUE DE LA PECHERIE DE CREVETTES DE LA BAIE DU REQUIN
Résumé
Etant donné que normalement la durée de vie exploitable des Peinaeidae ne dépasse pas un an environ, les captures pour une année donnée dépendent du nombre de recrues entrant dans la phase exploitable juste avant ou pendant cette année. Il est dono indispensable, aux fins de l'aménagement, de connaître les variations survenant dans l'époque et dans l'importance de ce recrutement.
Les modèles halieutiques étudiés reposent principalement sur les pêcheries de poissons de l'Atlantique Nord et ne conviennent donc pas aux pêcheries de Penaeidae de l'Australie-Occidentale. Les hypothèses de base sur le recrutement, l'estimation de l'âge, le schéma de croissance et les effets des migrations ne sont pas applicables.
Il est proposé un modèle reposant sur l'emploi de données détaillées sur les captures, l'effort de pêche et la composition par taille des prises. On y distingue entre les conséquences de la croissance, des migrations et des variations de densité avec le temps. Ce modèle fournit un moyen d'examiner l'époque, le lieu et l'importance du recrutement.
UN MODELO DESCRIPTIVO Y ANALITICO DE LA PESQUERIA DE CAMARONES DE LA BAHIA SHARK
Extracto
La duración de la vida pescable de los peneidos por lo general no es superior a un año aproximadamente y por esto la captura que pueda hacerse en un año determinado depende de los individuos que se incorporen a la pesquería exactamente antes de dicho año o durante él. El requisito básico para la ordenación es conocer las variaciones en cuanto al momento de verificarse el reclutamiento y a la intensidad del mismo.
Los modelos pesqueros examinados se basan primariamente en las pesquerías de peces del Atlántico norte, siendo inadecuados para las pesquerías de peneidos de Australia occidental. Son inaplicables los supuestos básicos sobre reclutamiento, estimación de la edad, tasa de crecimiento y efectos de la migración.
Se propone un modelo basado en el empleo de datos detallados sobre captura, esfuerzo y composición por tamaño de las capturas. Este modelo diferencia entre los efectos del crecimiento, la migración y la variación en densidad con el tiempo. Proporciona un medio para examinar la época, lugar e intensidad de los grupos de individuos que se incorporan a la pesquería.
Responses to changes in exploited populations are in general the result of a number of contributing responses, many not directly measurable. This makes a complete understanding of the systems involved extremely difficult. Models are simplified representations of these complex systems that (i) permit explanation of the observed phenomena, (ii) suggest the relationships between the significant parameters and (iii) provide a means of predicting changes that will occur within the system if the parameters are varied.
This approach is well known in fisheries science, and Beverton and Holt (1957) and Ricker (1948) have made fairly recent reviews of the literature. The models described by these workers have, however, been largely developed for teleost fisheries. Although the basic assumptions inherent in these models are upheld for prawn fisheries, certain statistics, particularly age, are difficult to determine.
Direct age determination of crustaceans in the catch is not possible and hence parameters such as age of recruitment (tr) and age of first capture (tc) from teleost models are not applicable. Size-age relationships are found only indirectly (Burken-road, 1951; Kurata, 1962). In addition, growth as measured by increases in external size is not continuous but takes place in a series of steps, each step occurring at moulting (Passano, 1960). Growth models commonly used (Von Bertalanffy, 1938; Parker and Larkin, 1959) are based on continuous growth, and these apply to crustacea only if size is assumed to increase during the intermoult. It has been found in most cases that these models empirically fit growth data, but no rigorous study has yet been made. Indeed, a form of step function may more adequately describe crustacean growth.
In teleost models, migration within the area under consideration is usually considered negligible, although Beverton and Holt (1957) propose a spawning migration model based on thermodynamic dispersal theory. A basic assumption of the population estimation model of DeLury (1947) is that migration does not occur. Penaeids are highly migratory (Weymouth, Lindner and Anderson, 1933; Lindner and Anderson, 1956; Dall, 1958; Slack-Smith, 1969) and migration is, therefore, an important parameter in penaeid populations.
A corollary to this high mobility is that recruitment must continue for an extended period if the numbers on a particular ground are to be maintained during the fishing season. Furthermore, in Shark Bay recruitment appears to be in waves, each probably associated with lunar periodicity in spawning and with later migration from prawn nursery areas (Racek, 1959; Dall, 1958).
Penaeid tagging for other than migration studies has not been entirely successful (Menzel, 1955; Dawson, 1957; Lindner and Anderson, 1956; Iversen and Idyll, 1960; Costello, 1959; Costello and Allen, 1960) although Lindner and Anderson (1956) recorded growth increments and Iversen (1962) made population estimates using Petersen tags.
In contrast to most penaeid fisheries, very detailed catch, effort and size composition data have been collected for the Shark Bay fishery (Slack-Smith, 1969; Slack-Smith and Stark, 1968). Insufficient research staff was available to prosecute a full biological programme, hence an attempt is made to use these commercial data to elucidate several biological processes within the fishery and to construct a model, initially descriptive but ultimately analytical, applicable to this population.
The general features of the fishery and of methods of data collection are described by Slack-Smith (1969) and Slack-Smith and Stark (1968).
Shark Bay penaeids seldom remain in the fishable stock for more than one season. The potential yield from an area, therefore, depends directly upon recruitment just prior to and during the fishing season. Catch and effort data from the Shark Bay fishery indicate that the time and place of recruitment throughout a particular season is not constant nor is it constant from season to season (Slack-Smith, 1966; 1969). A further complication is that the magnitude of total recruitment varies from year to year.
The basic management requirement for this fishery is to determine whether these changes are random, cyclic or trending downwards. If they are the result of a downward trend the causes of the decline must be determined and methods found by which it can be halted.
Before the nature of recruitment changes can be investigated, a means of estimating recruitment must be found. A model has accordingly been constructed to investigate these and other changes within the Shark Bay fishery.
The basic data for the proposed model are obtained from a computer programme which inputs scored length frequency data, the weight of the sample from which they are made, the total landed weight and the total fishing effort for a particular week (i). Numbers caught (Nijk) at each carapace length (j) are included in the output. This portion of the output is used to set up a three-dimensional array of Dijk (Fig. 1) that represents densities of prawns of particular sizes on each block (k) within the fishery during each week of the season, i.e.,
| Dijk = Nijk/fik | (1) |
The normal assumption that catch per unit of effort is proportional to absolute density is not entirely valid in the Shark Bay prawn fishery. Catchability, the constant relating these parameters, varies cyclically with changes in time of day and in lunar phase. It is assumed for this model that appropriate corrections have been made to catch rates to eliminate these effects.
To simplify explanation of this model, prawns are assumed to move on a line in the direction of migration, i.e., that the fishery is one block wide and there is no lateral emigration or immigration. In the case of the Shark Bay fishery this is not so and there is undoubtedly lateral movement, necessitating the inclusion of a two-dimensional migration component when the model is being tested on fishery data. Density changes within the model are more readily observed if each modal size group within a catch-per-length-frequency (there are usually 1 to 3) is represented by the modal value or values (MCLn). It is further assumed that these modal values always represent the densities of the whole group, i.e., there is no size selection during fishing, and the distribution around this mode remains constant.
As the season progresses, i.e., i increases (Fig. 1), each of these points of density will move through the space enclosed by the three axes of the model. The vector of this movement in i represents the time scale; the vector in j, size increase; the vector in k, migration.
2.1 Recruitment
Entry of a modal group into the model during the increase of i from o to λ (the length of the season) represents recruitment, and the associated j and k values give the modal recruit size and the area where juvenile prawns enter the fishery. The relative abundance of this recruit group is given by its MCLn value. The MCLn values are then summed to obtain an index of total recruitment for season s (Rs), i.e.
| Rs = ∑MCLn | (2) |
2.2 Growth and migration
Mean rate of size increase of a recruit group is represented by the rate at which the j component (growth increment) increases with time. Individual size increase is not continuous but is stepwise (section 1). However, in this model sudden increases in mean carapace length would be observed only if all prawns moulted at the same time. But, as moulting in Shark Bay prawns has been observed at virtually all times, it appears that in practice a continuous increase in mean size will be observed.
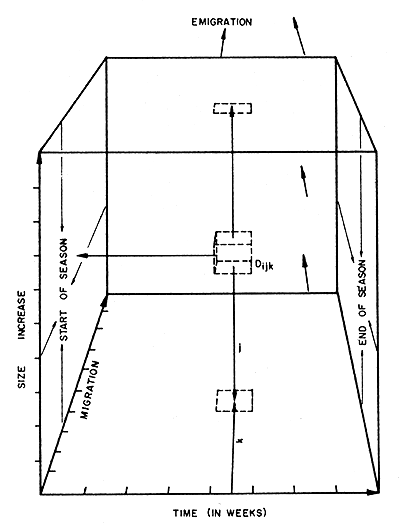
Fig. 1 Diagrammatic three-dimensional representation of the Shark Bay prawn fishery. Dijk is the density of a recruit group, MCLn, at time i, size j and position (after migration) k.
Migration rate (m) is given by the rate at which the k component increase with time.
2.3 Total mortality
The rate of density decrease in each recruit group is given by the rate of decrease of MCLn (Fig. 2). The Instantaneous Coefficient of Total Mortality (Z = F + M) can be calculated using methods described by Gulland (1965). If the abundance as given by MCLn is No at a given time to and Ni at time ti then
| Ni/No = S = e-Z (ti-to) | (3) |
i can be considered as a single week or any desired number of weeks. If the values for total mortality for a particular recruit group obtained from the model by equation (3) are plotted against the effort for that week, and provided the effort varies sufficiently, estimates of fishing mortality (F), natural mortality (M) and catchability (q) can be made for that group using the method described by Widrig (1954).
Prior to the construction of this model, components of migration were included in any estimate of Z made on a particular block in a similar manner, i.e.
| Z1 | = | F + M + E - I | (4) | |
| Where | Z1 | = | Instantaneous coefficient of total fall in numbers | |
| F | = | Instantaneous coefficient of fishing mortality | ||
| M | = | Instantaneous coefficient of natural mortality | ||
| I | = | Instantaneous coefficient of immigration | ||
| E | = | Instantaneous coefficient of emigration | ||
2.4 Emigration
Prawns lost from the fishable stock by emigration are represented by recruit groups passing through the rear face of the model. As each block in Shark Bay is a square of 10 × 10 mi, prawns migrating across the block at a rate m mi/wk will traverse m/10 of a block per wk. If the density on that block is Dijk, then the number of prawns emigrating from the fishery during that week is given by the product of that density and the area occupied by those prawns (m/10).
Fishing and natural mortalities also operate during that week, the numbers lost being represented by Dijk.m/10.(1-e-z)
If all these terms are summed for each week of the season the total emigration loss (E) is obtained, i.e.,
| E = ∑ Dijk.m/10. (1-e-z) | (5) |
2.5 Estimation of numbers
After entering the fishery the size of a particular recruit group (Rs) is decreased by the combined effects of natural and fishing mortality (Fig. 2), where Z = F + M. Now, Ni = No.e-(F + M)i, and if the contribution of M to total mortality is small the weekly rate of exploitation is approximated by (Ni-Ni-1)/Ni. The total number of prawns on the grounds at any particular week i (Nti) is given by
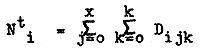
where k represents the number of blocks within the fishery.
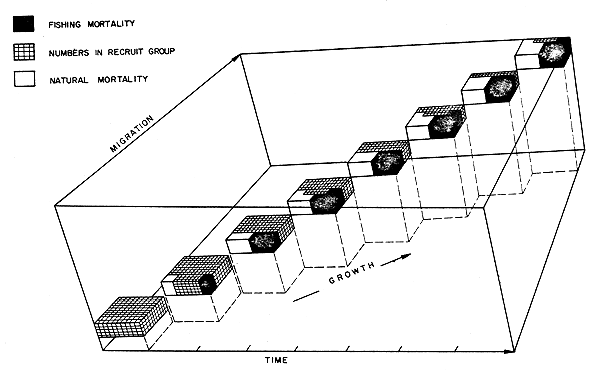
Fig. 2 Simplified representation of the fate of a recruit group, MCLn, after its entry into the Shark Bay fishery. At the end of each week the mean carapace length of the group increases by one unit and they migrate one unit. Fishing mortality (black) and natural mortality (white) are operating at the same rate. The number remaining at the end of each week is depicted by the grey shading. The co-ordinates of the model are given in Fig. 1.
Analytical aspects of the model require further development, and ultimately it is planned to programme estimation of the migration, growth amd mortality parameters as a single computer operation.
Two steps in the testing of these analytical aspects of the model will be used. The first is to simulate the model on the computer and, using iterative processes, estimate the most likely parameter values that produce results similar to those observed in the field. The second will be to make independent estimates of parameters and compare them with estimates obtained from the model. The best prospect of producing such independent estimates of growth, exploitation and migration is by tagging or marking. Estimates of numbers of juveniles leaving the nursery areas will be used for independent estimates of recruitment.
The relationship between catch per unit of effort (C/UE) and absolute density (D) is not simple. Cyclic changes in catchability associated with lunar and diurnal changes have already been postulated (Racek, 1959; Slack-Smith, 1969). Methods of making direct measurements of density are being explored so that these relationships can be investigated. Thus, catch and effort data can then be converted to actual density when exploitation and mortality rates and population size are being assessed.
The short life span of penaeids precludes any long term predictions of events based on biology or population dynamics within the fishery. There has been some success in associating population changes with the physico-chemical environment (St. Amant, Broome and Ford, 1966). However, the long-term prediction of these physico-chemical changes is notoriously difficult (Slack-Smith, 1969). It is considered, therefore, that the rather violent fluctuations in penaeid catch universally observed will remain largely unpredictable on a long-term basis.
If fishing intensity is sufficiently high to affect recruitment, then some form of short-term prediction of events is possible. However, Ingle (1961) states that it is most unlikely that fishing can ever be sufficiently intense to deplete spawners to the point of affecting recruitment, as prawn fecundity is extremely high.
Destruction of the biota and sediment balance by trawling in some areas (Slack-Smith, 1969) has been postulated as another possible cause of falling catch. However, rather than decreasing recruitment this would increase natural mortality or bring about changes in path or speed of migration.
Beverton, R.J.H. and S.J. Holt, 1957 On the dynamics of exploited fish populations. Fishery Invest., Lond.(2), 19:533 p.
Burkenroad, M.D., 1951 Measurement of natural growth rates of decapod crustaceans. Proc. Gulf Caribb.Fish.Inst., 3:25–6
Costello, T.J., 1959 Marking shrimp with biological stains. Proc.Gulf.Caribb.Fish.Inst., 11:1–6
Costello, T.J. and D.M. Allen, 1960 Notes on the migration and growth of pink shrimp (Penaeus duorarum). Proc.Gulf.Caribb.Fish.Inst., 12:5–9
Dall, W., 1958 Observations on the biology of the greentail prawn, Metapenaeus mastersii (Haswell) Crustacea Decapoda: Penaeidae). Aust.J.mar.Freshwat.Res., 9(1):111–34
Dawson, C.E., 1957 Studies on marking of commercial shrimp with biological stains. Spec. scient.Rep.U.S.Fish.Wild.Serv., (231):23 p.
DeLury, D.B., 1947 On the estimation of biological populations. Biometrics, 3(4):145–67
Gulland, J.A., 1965 Manual of methods for fish stock assessment. Part 1. Fish population analysis. FAO Fish.tech.Pap. (40), Rev.1:68 p.
Ingle, R.M., 1961 Synoptic rationale of existing Florida shrimp regulations. Proc.Gulf. Caribb.Fish.Inst., 13:22–7
Iversen, E.S., 1962 Estimating a population of shrimp by the use of catch per effort and tagging data. Bull.mar.Sci.Gulf.Caribb., 12(3):350–98
Iversen, E.S. and C.P. Idyll, 1960 Aspects of the biology of the Tortugas pink shrimp, Penaeus duorarum. Trans.Am.Fish.Soc., 89(1):1–8
Kurata, H., 1962 Studies on the age and growth of crustacea. Bull.Hokkaido reg.Fish.Res. Lab., (24):1–115
Lindner, M.J. and W.W. Anderson, 1956 Growth, migration, spawning and size distribution of shrimp Penaeus setiferus. Fishery Bull.Fish.Wild.Serv.U.S., (56):553–645
Menzel, R.W., 1955 Marking of shrimp. Science, 121(3143):446
Passano, L.M., 1960 Moulting and its control. In The physiology of Crustacea, edited by T.H. Waterman, London, Academic Press, pp. 473–536
Parker, R.R. and P.A. Larkin, 1959 A concept of growth, J.Fish.Res.Bd Can., 16(5):721–45
Racek, A.A., 1959 Prawn investigations in eastern Australia. Res.Bull.St.Fish.N.S.W., (6):1–57
Ricker, W.E., 1948 Methods of estimating vital statistics of fish populations. Indiana Univ.Publ.Sci.Ser., 15:1–101
Slack-Smith, R.J., 1966 The Shark Bay prawning industry. Western Australia, Government Printer, 13 p.
Slack-Smith, R.J., 1969 The prawn fishery of Shark Bay, Western Australia. FAO Fish. Rep., (57) Vol.3:717–34
Slack-Smith, R.J. and A.E. Stark, 1968 Automatic processing of Western Australian prawn fishermen's log books. FAO Fish.Rep., (57) Vol.2:309–19
St. Amant, L.S., 1966 J.G. Broom and T.B. Ford, Studies of the brown shrimp, Penaeus aztecus, in Barataria Bay, Louisiana, 1962–1965, Proc.Gulf Caribb.Fish.Inst. 18:1–17
Von Bertalanffy L., 1938 Inquiries into growth laws. 2. A quantative theory of organic growth. Hum.Biol., 10(2):181–213
Weymouth, F.M., M.J. 1933 Lindner and W.W. Anderson, Preliminary report on the life history of the Common Shrimp Penaeus setiferus (Linnaeus). Fishery Bull.Fish.Wildl. Serv.U.S., (48):1–26
Widrig, T.M., 1954 Methods of estimating fish populations with application to Pacific sardine. Fishery Bull.Fish Wild.Serv. U.S., (94):141–66
![]()
![]()
![]()