![]()
![]()
![]()
by
D. W. GOTSHALL
Marine Resources Operations
California Department of Fish and Game
Eureka, California, U. S. A.
Abstract
The food habits of fish were studied in conjunction with surveys of Pandalus jordani off northern California, to examine the predator-prey relationship between fish and ocean shrimp and to decide if a particular predator could be used to estimate relative abundance of ocean shrimp. After some study of the stomach contents of 16 species of fish, the Pacific hake, Merluccius productus, was chosen for detailed study.
Stomachs of 1,613 Pacific hake were examined between July 1964 and October 1966. The ocean shrimp year class composition in Pacific hake stomachs compared favorably with that estimated from sea surveys. Growth rates calculated for the youngest shrimp year class in the hake stomachs were similar to those calculated from market and sea survey samples. The average number of ocean shrimp in each year class per hake stomach was used to calculate annual mortality rates, which were quite consistent for the 1962, 1963 and 1964 year classes.
UTILISATION DES HABITUDES ALIMENTAIRES DES PREDATEURS POUR ESTIMER L'ABONDANCE RELATIVE DE LA CREVETTE Pandalus jordani RATHBUN
Résumé
On a examiné les habitudes alimentaires de certains poissons en les rapprochant d'études sur Pandalus jordani au large de la Californie du Nord, afin de voir, sur la base du rapport prédateur/proie existant entre poissons et cette crevette, si un certain prédateur pouvait servir à estimer l'abondance relative de P. jordani. Après analyse des contenus stomacaux de 16 espèces de poissons, c'est le merlu Merluccius pro-productus qui a été choisi pour étude plus approfondie.
Les estomacs de 1 613 merlus M. productus ont été examinés entre juillet 1964 et octobre 1966. La composition par classe d'âge des orevettes P. jordani trouvées dans les estomacs de M. productus cadrait bien avec les estimations fournies par des enquêtes en mer. Les taux de croissance calculés pour la plus jeune classe d'âge de crevettes trouvées dans les estomacs de merlus étaient analogues à ceux qui avaient été calculés sur la base d'échantillons provenant des marchés ou récoltés en mer. On a utilisé le nombre moyen de crevettes de chaque classe d'âge par estomac de merlu pour calculer les taux annuels de mortalité, qui se sont révélés très cohérents pour les classes 1962, 1963 et 1964.
EL EMPLEO DE LOS HABITOS ALIMENTARIOS DE LOS PREDADORES PARA ESTIMAR LA ABUNDANCIA RELATIVA DEL CAMARON, Pandalus jordani RATHBUN
Extracto
Se estudiaron los hábitos alimentarios de peces simultáneamente con reconocimientos de P. jordani en aguas del norte de California, para examinar la relación predadorpresa entre peces y camarones y poder decidir si un predador determinado podría emplearse para estimar la abundancia relativa del camarón. Después de examinar el contenido estomacal de 16 especies de peces, se seleccionó la merluza del Pacífico, Merluccius productus para realizar estudios detallados.
Entre julio de 1964 y octubre de 1966 se examinaron los estómagos de 1.613 merluzas del Pacífico. La composición por clases anuales del camarón en los estómagos de la merluza del Pacífico concordaba bastante bien con las estimaciones hechas a base de reconocimientos efectuados en el mar. Las tasas de crecimiento calculadas para la clase anual de camarón más joven encontrado en los estómagos de las merluzas eran análogas a las calculadas a partir de muestras obtenidas en los mercados y en el mar. Se empleó el número medio de camarones de cada clase anual por estómago de merluza para calcular las tasas de mortalidad anuales, que eran bastante constantes para las clases de 1962, 1963 y 1964.
A series of sea surveys was undertaken in the spring of 1965 to investigate California's northern ocean shrimp population, which has supported a commercial fishery since 1958 (Dahlstrom 1961). The purpose of the surveys was to obtain reliable estimates of population size, year class composition and mortality rates for use in developing a yield equation. The surveys were conducted in a 500 km2 area (Fig. 1) using a stratified random sampling design.
1.1 Predator food habit study
In conjunction with the sea surveys, we examined the stomach contents of larger fish, caught incidentally on the shrimp bed, to determine possible predator-prey relations. This study began in July 1964 when Pacific hake (Merluccius productus) were observed regurgitating large numbers of 1964 year class ocean shrimp (Pandalus jordani). We suspected that predator food habits might indicate the relative abundance of the incoming year class of shrimp and help us to follow their growth rates. Ocean shrimp do not become fully vulnerable to commercial nets until their second year of life, and even with a 1.27 cm stretch mesh liner in the cod end, significant numbers of younger shrimp escape.
1.2 Predators as biological samplers
The idea of using predators in population studies is not new. Thorson (1957) has suggested the use of predators to estimate benthic productivity. From stomach contents of Atlantic cod (Gadus callarias) Powles (1965) estimated the mortality of plaice (Hippoglossoides platessoides) due to Atlantic cod grazing. Gotshall, Smith and Holbert (1965) suggested that blue rookfish (Sebastodes mystinus) stomach contents may be used to indicate relative abundance of juvenile and larval crustaceans and fish.
2.1 Collection and examination of stomachs
Stomachs were obtained from fish caught on the shrimp bed (Fig. 1) in various types of otter trawls, including those of commercial shrimp vessels (3.81 cm mesh), bottomfish trawlers (11.43 cm mesh), and California Department of Fish and Game research vessels. On the sea surveys, research vessels used a 12.50 m semi-balloon Gulf shrimp trawl with 2.54 cm stretch mesh and 1.27 cm stretch mesh liner in the cod end. Stomachs were removed from the fish soon after capture and preserved in 70 per cent isopropyl alcohol. When the stomachs were removed, organisms found in the mouths and throats of the fishes were discarded. Data collected from each fish included: species, total length, sex, date and location of capture.
Stomach contents were examined under a dissecting microscope. Identifiable organisms were measured, counted, and their volumetric displacement determined in a graduated cylinder.
2.2 Ocean shrimp age determination
Ocean shrimp are aged in California by a combination of criteria using carapace length frequencies. These include growth rate studies and age of sexual change. California studies indicate that all male ocean shrimp enter the transitional phase before they are two years old, and all transitionals become females before their third year of life (Dahlstrom, in press). Studies by Butler (1964) have shown this to be true also with British Columbia ocean shrimp.
Fig. 1 Area in which the surveys were conducted.
One and two-year-old shrimps are separated at that size immediately following the last male measurement where the frequency distribution reaches the minimum. Two and three-year-old shrimps are divided at the minimum frequency immediately following the last transitional measure. For example, in August 1966 the 1965 year class of ocean shrimps captured off northern California had a size range of 14.4 to 18.6 mm, 1964 year class shrimp ranged in size from 18.8 to 22.6 mm and 1963 year class shrimp ranged from 22.8 to 24.2 mm carapace length.
2.3 Population estimates from sea survey
The stratified random sampling program used on our sea survey was designed by our biostatistical staff. A manuscript describing the method is in preparation (Abramson, pers. comm.) We are also in the process of preparing a paper describing the results of the sea surveys. The survey area (Fig. 1) was divided into 14 strata of unequal size. The first 10 strata made up the area of consistent commercial fishing success, based on catch data dating back to 1952. Strata 11 through 14 consisted of areas of limited commercial fishing success. Thirty-nine randomly selected trawls were assigned to the survey area (excluding known snag areas). If these were completed, an additional 39 trawls were attempted, and so on until the number of days allotted for the survey were used up. Ten of the 39 trawls were assigned to strata numbers 11 through 14, the number per stratum depending upon the area of the stratum. The remaining 29 trawls were assigned to the first 10 strata, the number per stratum again depending on the area.
The length of each trawl is 0.8 km, and the net opening averages 7.6 m; thus the area swept by the trawl equals 6131.6 m2.
The shrimps from each trawl were placed in sampling boxes, each box containing 80 equal sized compartments. Two samples were selected from the boxes using random number lists. The shrimps in the samples were measured, sexed and weighed to determine the age composition and total weight of the catch. All of the catch data were processed by computer (Abramson, pers. comm.). The resulting mean catch per trawl for each stratum was expanded to an estimated total population for the stratum by multiplying it by the total number of possible trawls in the strata. The estimated population of each year class is equal to the sum of the estimates in the 14 strata.
2.4 Market sampling
Samples of ocean shrimp are obtained from northern California commercial landings through the season at Eureka and Crescent City, and as many of the landings as possible are sampled. The sample consists of filling a one liter container with shrimp from the catch. The sample may range from as few as 50 shrimps to over 150 shrimps, depending on the number of one-year-old shrimps in the sample. During the 1966 season (May to 7 August) a total of 15 boats made 363 landings. A total of 155 samples containing 17,474 shrimps were obtained from these landings.
3.1 July to September 1964 study
We examined 242 stomachs from 16 species of fish, collected from July to September, 1964. Stomach contents were analyzed in some detail but only results relating to ocean shrimp are included in the present paper. Ocean shrimps were found in stomachs from 8 species of fish. Pacific hake (Merluccius productus) and arrowtooth flounder (Atheresthes stomias) were selected for a one year intensive analysis of stomach contents. These two species were chosen because they were easy to obtain and a large proportion of their stomachs contained ocean shrimp; 97 of 164 hake stomachs, and 10 of 15 flounder stomachs.
Other species from which a large proportion of stomachs contained ocean shrimp were flat rockfish (Sebastodes rubrivinctus), sablefish (Anoplopoma fimbria), petrale sole (Eopsetta jordani), yellowtail rockfish (S. flavidus), greenstripe rockfish (S. elongatus), and stripetail rockfish (S. saxicola). These fish were not included in the 12-month study because large numbers of fish were difficult to obtain, or, in the case of the rockfish, because of the tendency for the expanding swim bladder to evert the stomach when brought to the surface from depths. It is unfortunate that flag rockfish could not be used, because the two stomachs of this species examined contained 170 juvenile (1964 year class) and adult ocean shrimp.
3.2 Arrowtooth flounder study
Arrowtooth flounder stomachs were collected monthly from September 1964 to October 1965, except in January 1965. We examined 425 stomachs during this period and found that 172 were empty. Juvenile shrimps (1964 and 1965 year classes) were present in 33 stomachs. Adult shrimps (1962, 1963 and 1964 year classes) were found in 102 stomachs, but were not present in those collected during November and December 1964 or February and March 1965. This is not surprising since the December 1964 and February 1965 stomachs were not from fish captured on the shrimp bed. The lack of shrimps in the November and March stomachs was probably due to the small sample. The stomachs from December collections were from 185 fathoms (340 m) west of Eureka, California, and the February samples were trawled west of Patricks Point in 380 fathoms (660 m). Crustaceans as a group were the dominant food of arrowtooth flounder during the study period.
3.3 Pacific hake study
A total of 527 Pacific hake stomachs was examined from July 1964 to September 1965 and 77 were empty. Adult shrimp (1961, 1962, 1963 and 1964 year classes) were found in 202 stomachs and juvenile shrimp (1964 and 1965 year classes) in 131 of the stomachs. Euphausiids were present in more stomachs than any other organism. Slender sole Lyopsetta exilis was the dominant food fish, appearing in 17 stomachs. These flatfish were also the fish most frequently taken in our survey trawls.
When we compared the number of ocean shrimp per arrowtooth flounder stomach and Pacific hake stomach, we found that Pacific hake not only contained more shrimp per stomach, but had preyed upon all of the year classes present in the shrimp population, including the outgoing 1961 year class which was not represented in the flounder stomachs. For these reasons Pacific hake were selected as our biological samplers and we discontinued collecting stomachs from flounder.
3.4 Pacific hake study, October 1965/66
Beginning in October 1965 the method of analysis of stomach contents was modified. Instead of preserving the stomachs, their contents were removed and examined at the time of collection, and organisms other than ocean shrimp were discarded. After October 1965, we examined 1,086 Pacific hake stomachs, of which 54 were empty.
3.4.1 Relative abundance of ocean shrimp year classes in Pacific hake stomachs
The 1964 year class was the most numerous age group found in hake stomachs from July 1964 to July 1966 (Table I). This year class provided the greatest portion of the catch of the commercial fishery during the 1965 and 1966 seasons. Our sea surveys indicated that the 1964 year class dominated the northern California ocean shrimp population until the spring of 1966 (Table II).
The 1965 year class was represented by fewer shrimp in both the hake stomachs and the fishery. Our sea surveys confirmed that this year class was not as numerous as the 1964 year class. Both Pacific hake stomachs and the sea survey indicated that the incoming 1966 year class was more abundant than the 1965 year class, but not as abundant as the 1964 year class (Tables I and II).
TABLE I
Numbers of 1961, 1962, 1963, 1964, 1965 and 1966 year class ocean shrimp per Pacific hake stomach collected off Northern California 1, July 1964 2 - October 1966
| Year Class | No. of stomachs 3 | No. of cean shrimp | ||||||
| 1961 | 1962 | 1963 | 1964 | 1965 | 1966 | |||
| 1964 | ||||||||
July | 0.44 | 1.38 | 16 | 29 | ||||
Aug. | 0.03 | 0.45 | 1.23 | 1.44 | 75 | 236 | ||
Sept. | 0.07 | 0.28 | 5.05 | 42 | 227 | |||
Oct. | 0.01 | 0.04 | 0.03 | 8.10 | 73 | 597 | ||
| 1965 | ||||||||
March | 0.66 | 3 | 2 | |||||
April | 0.06 | 0.07 | 0.72 | 69 | 59 | |||
May | 0.13 | 0.13 | 2.10 | 30 | 71 | |||
June | 0.49 | 43 | 21 | |||||
July | 0.09 | 2.85 | 0.09 | 54 | 164 | |||
Aug. | 0.07 | 0.37 | 1.59 | 0.78 | 27 | 76 | ||
Sept. | 0.50 | 1.25 | 0.08 | 12 | 22 | |||
Oct. | 0.01 | 0.08 | 1.45 | 0.39 | 152 | 291 | ||
| 1966 | ||||||||
April | 0.05 | 0.20 | 0.43 | 210 | 135 | |||
May | 0.11 | 1.78 | 0.67 | 9 | 23 | |||
June | 0.03 | 0.85 | 0.42 | 0.08 | 190 | 262 | ||
July | 0.77 | 0.38 | 0.08 | 74 | 91 | |||
Aug. | 0.10 | 0.18 | 0.46 | 150 | 111 | |||
Sept. | 0.52 | 0.61 | 2.60 | 122 | 455 | |||
Oct. | 0.08 | 0.75 | 1.58 | 5.84 | 125 | 1 020 | ||
1 107 September and all October 1966 stomachs collected off Brookings, Oregon
2 December 1964 not included because hake were not captured on shrimp bed
3 Does not include empty stomachs
TABLE II
Numbers of 1962, 1963, 1964, 1965 and 1966 year class shrimp in population surveyed off Northern California and Southern Oregon, October 1964 – October 1966
| Survey Period | Area | Year Class (Millions) | |||||
| 1961 | 1962 | 1963 | 1964 | 1965 | 1966 | ||
| 6–12 Sept. 1964 | Cape Ferralo to Redding Rock | 6 | 51 | 86 | 1,038 | ||
| 10 Mar.-23 Apr. 1965 | Pelican Bay1 Mad River | 14 | 76 | 797 | |||
| 1 June-1 July 1965 | " " | 6 | 64 | 961 | |||
| 27 Sept.-22 Oct. 1965 | " " | 6 | 59 | 536 | 76 | ||
| 18 Mar.-27 Apr. 1966 | " " | 20 | 372 | 355 | |||
| 15 Aug.-10 Sept. 1966 2 | " " | 3 | 156 | 183 | 1,766 | ||
1 See Fig. 1
2 Includes estimate of population off Brookings, Oregon, made by Oregon Fish Commission
In comparing the percentage composition of the various year classes of shrimp in hake stomachs with our sea survey data and commercial catch samples (Table III), we find that year class compositions determined from stomach and sea surveys are quite similar, while hake stomach results differ from those obtained from commercial landings. This is what we would expect if the food intake of hake is a representative sample of the shrimp population. We assume that our sea surveys reliably measure the relative abundance of all but the incoming year class. In the autumn, the stomachs appear to yield a more accurate index of the abundance for the incoming year class than do our sea surveys.
3.4.2 Length frequencies of incoming year class Assuming Pacific hake are randomly feeding on ocean shrimp, the length composition of shrimp from stomachs should reflect the size composition of the population. To illustrate this relation, we can compare shrimp lengths of incoming year classes obtained from various sources, since there is no overlap with preceding year class lengths.
Comparative size data are available for the 1964, 1965 and 1966 incoming year classes (Fig. 2). The length frequency modes from the stomachs and commercial or sea survey samples are the same in each case except in August 1966. Generally, Pacific hake stomachs yielded a wider range of sizes and tended to include smaller shrimp than either the sea surveys or commercial fishery samples. From these comparisons it would seem that ocean shrimp growth rates could be followed by using hake as samplers.
3.4.3 Mortality rates Another method for comparing the effectiveness of Pacific hake as biological samplers is to examine mortality rates calculated from the numbers of shrimp in hake stomachs and sea survey data.
The following equation was used to calculate total mortality rates (Ricker, 1958).
it = - loge (Nt/No)
where:
t = the fraction of the year between the estimate of No and Nt
i = instantaneous rate of total mortality
loge = natural or naperian logarithms
Nt = index or numbers of a given year class at time t
No = index or numbers of a given year class at some time previous to t
Annual total mortality rates (a) were derived from Ricker's tables. Values of N for hake stomach data for autumn months were calculated by adding together the number of shrimp of a given year class for July through October and dividing by the total number of stomachs (excluding empty stomachs). The spring N value was based on the mean number of shrimp per stomach from April through July (Tables IV, V and VI). The sea survey estimates of N are the result of our stratified random sampling except for the September 1964 survey (Table II). This survey was conducted using random and non-random tows over a smaller sampling area. We believe, therefore, that the 1964 survey produced under-estimates of population size and of mortality rates.
The autumn 1966 population estimates include the population off Brookings, Oregon. These estimates are the result of an October sea survey conducted by the Oregon Fish Commission that used a sampling method similar to ours (Robinson, pers. comm.). The population off Brookings consisted of 1,430 million 1966 year class shrimp, 67 million 1965 year class shrimp, and 64 million 1964 year class shrimp. They were included because we have reason to believe that these shrimp are part of the northern California population that moved with a subsurface current into Oregon waters.
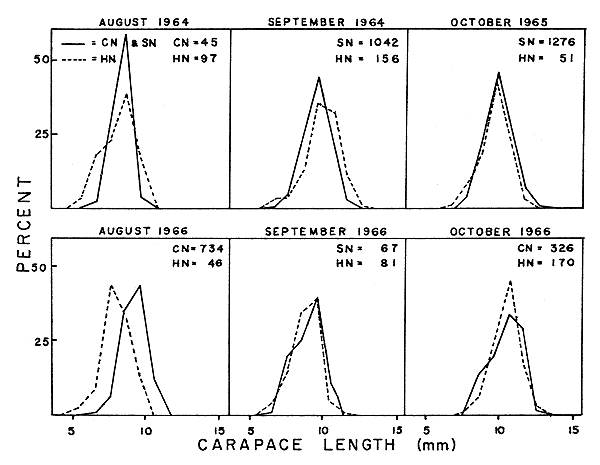
CN = NUMBER OF SHRIMP FROM COMMERCIAL SAMPLES
SN = NUMBER OF SHRIMP FROM SEA SURVEY SAMPLES
HN = NUMBER OF SHRIMP FROM HAKE STOMACHS
Fig. 2 Comparative size data for the 1964, 1965 and 1966 incoming year classes.
TABLE III
Comparison 1 of percent composition (by numbers) of Northern California ocean shrimp year classes in Pacific hake stomachs with fishery (market) sampling and sea surveys of population, July 1964–October 1966
| Year Class | ||||||
| 1961 | 1962 | 1963 | 1964 | 1965 | 1966 | |
| 1964 | ||||||
July | ||||||
Hake | 24.1 | 75.9 | ||||
Fishery | 2.1 | 47.4 | 50.5 | |||
August | ||||||
Hake | 0.8 | 14.4 | 39.0 | 45.8 | ||
Fishery | 2.1 | 43.1 | 54.8 | |||
September | ||||||
Hake | 1.3 | 5.3 | 93.4 | |||
Sea survey | 0.6 | 5.2 | 6.2 | 88.0 | ||
| 1965 | ||||||
April | ||||||
Hake | 9.8 | 8.2 | 82.0 | |||
Sea survey | 3.4 | 10.8 | 85.8 | |||
May | ||||||
Hake | 5.6 | 5.6 | 88.7 | |||
Fishery | 11.3 | 27.5 | 61.2 | |||
June | ||||||
Hake | 100.0 | |||||
Fishery | 2.9 | 26.5 | 70.6 | |||
Sea survey | 0.6 | 6.2 | 93.2 | |||
July | ||||||
Hake | 3.0 | 94.0 | 3.0 | |||
Fishery | 1.3 | 11.9 | 86.8 | |||
October | ||||||
Hake | 0.3 | 4.1 | 74.9 | 20.6 | ||
Sea survey | 1.1 | 8.7 | 78.7 | 11.6 | ||
| 1966 | ||||||
April | ||||||
Hake | 2.7 | 49.8 | 47.5 | |||
Sea survey | 0.7 | 31.8 | 67.4 | |||
May | ||||||
Hake | 4.3 | 69.6 | 26.1 | |||
Fishery | 5.8 | 79.5 | 14.7 | |||
June | ||||||
Hake | 2.3 | 61.4 | 30.2 | 6.1 | ||
Fishery | 3.5 | 73.5 | 23.0 | |||
July | ||||||
Hake | 64.0 | 29.2 | 6.7 | |||
Fishery | 3.1 | 71.7 | 25.2 | |||
August | ||||||
Hake | 13.5 | 24.3 | 62.2 | |||
Fishery | 3.1 | 69.9 | 26.8 | 0.2 | ||
Sea survey | 0.5 | 13.8 | 22.1 | 63.6 | ||
September2 | ||||||
Hake | 14.2 | 16.2 | 69.5 | |||
Fishery | 0.2 | 55.7 | 39.1 | 5.0 | ||
October | ||||||
Hake | 0.1 | 9.2 | 19.3 | 71.4 | ||
Fishery | 0.4 | 51.0 | 34.8 | 13.8 | ||
Sea survey 3 | 4.1 | 4.3 | 91.6 | |||
1 Comparative fishery or sea survey data not available during October 1964 or August and September 1965
2 Pacific hake and commercial fishery data from Brookings, Oregon
3 Conducted by Oregon Fish Commission (Jack Robinson pers. comm.)
TABLE IV
Comparison of instantaneous mortality rates and annual mortality rates of 1962 year class Northern California ocean shrimp calculated from Pacific hake stomachs 1 and sea surveys 2 1964 – 1965
| Nt | No | it | t | i | a | |
| Autumn 1964 to Spring 1965 | ||||||
Hake | .05 | .19 | 1.34 | 0.750 | 1.78 | 0.83 |
Sea survey 3 | 14 | 51 | 1.29 | 0.583 | 2.21 | 0.89 |
Sea survey 4 | 6 | 51 | 2.14 | 0.750 | 2.85 | 0.94 |
| Autumn 1964 to Autumn 1965 | ||||||
Hake | .01 | .19 | 2.94 | 1.000 | 2.94 | 0.95 |
Sea survey | 8 | 51 | 1.85 | 1.083 | 1.70 | 0.82 |
| Spring 1965 to Autumn 1965 | ||||||
Hake | .01 | .05 | 1.609 | 0.333 | 4.83 | 1.00 |
Sea survey 3 | 8 | 14 | 0.560 | 0.5000 | 1.12 | 0.67 |
Sea survey 4 | 8 | 6 | -- | -- | -- | -- |
1 Spring value calculated from total shrimp divided by total stomachs collected from April through July, Autumn value, equals total shrimp divided by total stomachs collected from July through October
2 Survey dates: September 1964, April 1965, June 1965, October 1965, April 1966, and August 1966
3 Survey, April 1965
4 Survey, June 1965
TABLE V
Comparison of instantaneous mortality rates and annual mortality rates of 1963 year class Northern California ocean shrimp calculated from Pacific hake stomachs 1 and sea surveys 2 1964 – 1966
| Nt | No | it | t | i | a | |
| Autumn 1964 to Spring 1965 | ||||||
Hake | 0.07 | 0.55 | 1.976 | 0.750 | 2.63 | 0.93 |
Sea survey 3 | 76 | 86 | .124 | 0.583 | 0.21 | 0.20 |
Sea survey 4 | 64 | 86 | .295 | 0.750 | 0.39 | 0.32 |
| Autumn 1964 to Autumn 1965 | ||||||
Hake | 0.14 | 0.55 | 1.283 | 1.000 | 1.28 | 0.72 |
Sea survey | 59 | 86 | .377 | 1.083 | 0.35 | 0.30 |
| Spring 1965 to Autumn 1965 | ||||||
Hake | 0.14 | .07 | ||||
Sea survey 3 | 59 | 76 | .253 | 0.500 | 0.51 | 0.40 |
Sea survey 4 | 59 | 64 | .081 | 0.333 | 0.24 | 0.21 |
| Spring 1965 to Spring 1966 | ||||||
Hake | 0.02 | 0.07 | 1.253 | 1.000 | 1.25 | 0.71 |
Sea survey 3 | 20 | 76 | 1.335 | 1.000 | 1.34 | 0.74 |
Sea survey 4 | 20 | 64 | 1.163 | 0.833 | 1.40 | 0.75 |
| Autumn 1965 to Spring 1966 | ||||||
Hake | 0.02 | 0.14 | 1.946 | 0.750 | 2.59 | 0.92 |
Sea survey | 20 | 59 | 1.082 | 0.500 | 2.16 | 0.88 |
| Autumn 1965 to Autumn 1966 | ||||||
Hake | 0.002 | 0.14 | 4.248 | 1.000 | 4.25 | 0.99 |
Sea survey | 3 | 59 | 2.979 | 0.833 | 3.58 | 0.97 |
| Spring 1966 to Autumn 1966 | ||||||
Hake | 0.002 | 0.02 | 2.302 | 0.333 | 6.91 | 1.00 |
Sea survey | 3 | 20 | 1.897 | 0.333 | 5.69 | 1.00 |
1 Spring value calculated from total shrimp divided by total stomachs collected from April through July, Autumn value, equals total shrimp divided by total stomachs collected from July through October
2 Survey dates: September 1964, April 1965, June 1965, October 1965, April 1966, and August 1966
3 Survey, April 1965
4 Survey, June 1965
The 1965 year class was not included in the comparisons because the autumn 1965 mean N was smaller than the spring and autumn 1965 mean N values.
3.4.3.1 Year class mortality The a values from the sea surveys and stomachs for the 1962 year class are quite similar (Table IV). The a value from hake stomachs increased with time until it reached the maximum value of 1.00 between the spring and autumn of 1965. This is expected for this age class, as it apparently suffers total mortality during the winter and spring of the fourth year of life (Dahlstrom, in press; Butler, 1964).
Annual mortality rates for the 1963 and 1964 year class from the shrimp per stomach and sea survey data did not agree as closely as those calculated for the 1962 year class (Tables V and VI). Best agreement was from the spring of 1965 to the spring of 1966 and autumn of 1965 to autumn of 1966. The greatest discrepancy in estimates occurred from the autumn of 1964 to spring 1965.
Generally the annual mortality rates derived from Pacific hake stomachs were higher than those calculated from the sea surveys. The small time intervals (t) for the shrimp per stomach annual mortality rate generally yielded lower estimates of a. Total annual mortality (a) could not be calculated for the 1963 or 1964 year class from the spring 1965 to autumn 1965 N values.
The most significant a values in terms of consistency were those derived from the hake stomachs from autumn 1964 to spring 1965, and autumn 1965 to spring 1966. The 1963 year class a values for these periods were 0.93 and 0.92 respectively, and the 1964 year class estimates were 0.78 and 0.78. Since this mortality took place when there was no fishery, the annual mortality rate (a) is equivalent to the annual natural mortality rate (n) (Ricker, 1958).
The concept of predator sampling involves two basic assumptions:
That the predator randomly feeds on the prey species throughout its range and does not select a particular size or age
That all sizes or ages of the species under study are equally vulnerable to the predator
We must consider several possible sources of error in using predators as biological samplers. One of the first questions that arose in our study was: are the Pacific hake or other predators feeding on the shrimps that are in the net? We have no doubt that a few shrimps find their way into the mouths and throats, or even as far as the stomach. However, if all or most of the shrimps found in the stomachs were picked up in the net, then hake collected with the commercial bottomfish trawls (11.43 cm mesh) would not contain nearly as many shrimps as those caught with the shrimp trawls (3.81 cm mesh). The highest number of shrimp per stomach occurred during October 1964 when all the hake sampled were obtained from commercial bottomfish trawlers (Table I). Also, there was a difference between the age composition of shrimps in hake stomachs and the age composition of the commercial shrimp fishery samples, and close agreement between age composition of shrimps in hake stomachs and those in sea survey samples (Table III). This would be expected if Pacific hake feed on shrimps outside the net.
TABLE VI
Comparison of instantaneous mortality rates and annual mortality rates of 1964 year class Northern California ocean shrimp calculated from Pacific hake stomachs 1 and sea survey 2
| Nt | No | it | t | i | a | |
| Autumn 1964 to Spring 1965 | ||||||
Hake | 1.47 | 4.53 | 1.125 | 0.750 | 1.50 | .78 |
Sea survey 3 | 797 | 1038 | 0.266 | 0.583 | 0.46 | .37 |
Sea survey 4 | 961 | 1038 | 0.079 | 0.750 | 0.10 | .10 |
| Autumn 1964 to Autumn 1965 | ||||||
Hake | 1.76 | 4.53 | .945 | 1.000 | 0.94 | .61 |
Sea survey | 536 | 1038 | 0.663 | 1.083 | 0.61 | .46 |
| Spring 1965 to Autumn 1965 | ||||||
Hake | 1.76 | 1.47 | ||||
Sea survey 3 | 536 | 797 | 0.397 | 0.500 | 0.79 | .55 |
Sea survey 4 | 536 | 961 | 0.584 | 0.333 | 1.75 | .83 |
| Spring 1965 to Spring 1966 | ||||||
Hake | 0.57 | 1.47 | 0.947 | 1.000 | 0.95 | .61 |
Sea survey 3 | 372 | 797 | 0.762 | 1.000 | 0.76 | .53 |
Sea survey 4 | 372 | 961 | 0.949 | 0.833 | 1.14 | .68 |
| Autumn 1965 to Spring 1966 | ||||||
Hake | 0.57 | 1.76 | 1.127 | 0.750 | 1.50 | .78 |
Sea survey | 372 | 536 | .365 | 0.500 | 0.73 | .52 |
| Autumn 1965 to Autumn 1966 | ||||||
Hake | 0.49 | 1.76 | 1.279 | 1.000 | 1.28 | .72 |
Sea survey | 156 5 | 536 | 1.234 | 0.833 | 1.48 | .77 |
| Spring 1966 to Autumn 1966 | ||||||
Hake | 0.49 | 0.57 | 0.151 | 0.333 | 0.45 | .36 |
Sea survey | 156 | 372 | .869 | 0.333 | 2.61 | .93 |
1 Spring value calculated from total shrimp divided by total stomachs collected from April through July. Autumn value, equals total shrimp divided by total stomachs collected from July through October
2 Survey dates: September 1964, April 1965, June 1965, October 1965, April 1966, and August 1966
3 Survey, April 1965
4 Survey, June 1965
5 Includes population off Brookings, Oregon
TABLE VII
Comparison of number of ocean shrimp found in stomachs from various sizes of Pacific hake, July 1964–October 1966
| Total Length Groups (cm) | ||||||||
| 10–19 | 20–29 | 30–39 | 40–49 | 50–59 | 60–69 | 70–79 | 80–89 | |
| July-October 1964 | ||||||||
Number of stomachs 1 | 4 | 11 | 6 | 62 | 67 | 52 | 3 | 0 |
Number without shrimp | 4 | 10 | 0 | 14 | 12 | 17 | 0 | 0 |
Number of shrimp | 0 | 1 | 12 | 113 | 527 | 415 | 13 | 0 |
Shrimp per stomach | 0.00 | 0.09 | 2.00 | 1.82 | 7.86 | 7.98 | 4.33 | 0.00 |
| March-October 1965 | ||||||||
Number of stomachs 1 | 12 | 10 | 186 | 152 | 38 | 13 | 0 | |
Number without shrimp | 8 | 7 | 91 | 65 | 13 | 4 | 0 | |
Number of shrimp | 4 | 4 | 235 | 299 | 122 | 38 | 0 | |
Shrimp per stomach | .33 | .40 | 1.26 | 1.97 | 3.21 | 2.92 | 0.00 | |
| April-October 1966 | ||||||||
Number of stomachs 1 | 12 | 35 | 481 | 238 | 92 | 8 | 1 | |
Number without shrimp | 12 | 25 | 234 | 95 | 29 | 3 | 0 | |
Number of shrimp | 0 | 15 | 521 | 696 | 734 | 60 | 11 | |
Shrimp per stomach | 0 | .43 | 1.08 | 2.92 | 7.98 | 7.50 | 11.00 | |
1 Does not include empty stomachs
The small samples of hake stomachs during months when Pacific hake were scarce did not contain shrimp of all year classes in the fishery (Table I). One method of increasing the number of shrimp per stomach in our samples would be to consider only the larger hake, but a casual analysis of the size composition of shrimp found in stomachs from various sizes of hake suggests that larger hake tend to feed more on the smaller shrimp. Pacific hake 20.0 to 49.9 cm TL were compared with hake 50.0 to 79.9 cm TL; 65 percent of the shrimp in the stomachs from the smaller size group of hake were juveniles, as compared with 87 percent in the stomachs from the larger hake. Pacific hake 50 cm or longer in total length had from two to three times as many shrimps per stomach as the smaller hake (Table VII).
From the sea surveys, we know ocean shrimp year classes are not randomly distributed over the bed, therefore an attempt was made to obtain hake from throughout the shrimp bed. Nevertheless, some of the wide variation in our shrimp per stomach data may be attributed to this bias.
The relative abundance of Pacific hake in the shrimp area fluctuates widely during the year. They are usually absent from late November through March, and start appearing in the nets during April with the largest catches occurring during July, August and September. Unfortunately, from the data collected we cannot evaluate the effect of changes in hake abundance on hake predation of shrimp. It is interesting to note, however, that the highest shrimp per stomach values occurred during autumn, when the frequency of hake in survey tows was highest (Table I).
The reason for what appears to be an underestimate of the 1965 year class in hake stomachs in the autumn of 1965 and spring of 1966 remains a mystery, although we suspect that the smallness of the hake samples and bias from catch locations are probably both involved.
The entire concept of predator sampling requires further study. Future plans for research on the hake/shrimp relationship include designing a statistically valid sampling program, and further analysis of data to determine the best method for estimating various mortality rates.
Eventually it may be possible to discontinue our costly sea surveys, and derive annual yield estimates from predator stomach data and commercial fishing trends. We feel the concept of predator sampling has potential as a fisheries management tool. It remains for fishery biologists to develop the method.
Butler, T.H. 1964 Growth, reproduction, and distribution of pandalid shrimps in British Columbia. J.Fish.Res.Bd Can., 21(6):1403–52
Dahlstrom, W.A., 1961 The California ocean shrimp fishery. Bull.Pacif.mar.Fish.Commn, (5):17–23
Dahlstrom, W.A. (in press) Synopsis of biological data on ocean shrimp, Pandalus jordani Rathbun. Paper presented to the FAO World Scientific Conference on the Biology and Culture of Shrimps and Prawns, Mexico, 12–21 June, 1967
Gotshall, D.W., J.G. Smith and A. Holbert, 1965 Food of the blue rockfish Sebastodes mystinus. Calif. Fish Game, 51(3):147–62
Powles, P.M., 1965 Life history and ecology of American plaice (Hippoglosoides platessoides F.) in the Magdalen Shallows. J.Fish.Res.Bd. Can., 22(1):565–98
Ricker, W.E., 1958 Handbook of computations for biological statistics of fish populations. Bull.Fish.Res.Bd Can., (119):300
Thorson, G., 1957 Bottom communities (Sublittoral or shallow shelf). In Treatise on marine ecology and paleoecology, edited by J.W. Hedgpeth. Mem.geol.Soc.Am. 67(1):461–534
Acknowledgments
This study would not have been possible without the assistance and cooperation of the following: Kim Ujita, Dave Hoopaugh, Ron Rinaldo, Gary Varney, Bob Jubenville, Steve Taylor, and Mel Willis who assisted in collecting and examining stomachs; Mel Willis prepared the figures; John Fitch, Bob Lavenberg and Dick Pool identified many of the fish and crustaceans found in the stomachs; Jack Robinson supplied the Oregon shrimp survey data; and Arlene Hardwick typed the original manuscript. I would also like to acknowledge the help of numerous commercial fishermen and the crew of the research vessel, N.B. Scofield, for their assistance in providing the fish involved in this study.
![]()
![]()
![]()