![]()
![]()
![]()
par
HENRI HOESTLANDT
Laboratoire de Zoologie
de l'Université Catholique de Lyon
Lyon, Rhône, France
Résumé
Cette étude constitue une première recherche sur le cycle de la crevette Penaeus duorarum Burkenroad, en Afrique Equatoriale (Dahomey); elle a eu lieu d'octobre 1962 à avril 1964.
Ce sont les post-larves qui migrent de la mer vers les eaux saumâtres lagunaires. La relation entre la salinité et la densité de quatre populations ainsi que la croissance de ces populations sont étudiées mensuellement, ce qui entraîne l'examen de 10 000 P. duorarum. Les jeunes crevettes ne peuvent vivre en eau douce; elles supportent des salinités faibles de 0,5‰ mais ne se développent normalement qu'à des salinités de 5 à 34‰. L'alimentation est principalement d'origine animale (petits poissons, crustacés, polychètes).
Après quelques mois de vie en eau saumâtre, les crevettes se dirigent vers la mer et sont principalement pêchées sur les fonds vaseux de 40 à 55 m.
Aux divers stades (post-larves, jeunes crevettes, crevettes adultes), les crevettes Penaeus duorarum du Dahomey sont un peu plus grandes que celles de la Floride.
RESEARCH ON THE BIOLOGICAL CYCLE OF Penaeus duorarum BURKENROAD 1939 IN DAHOMEY
Abstract
This paper constitutes initial research on the life cycle of Penaeus duorarum Burkenroad, in Equatorial Africa (Dahomey), lasting from October 1962 to April 1964.
The shrimps migrate at the post-larval stage from the sea in the direction of the brackish waters of the coastal lagoons. The relationships between salinity and the density of four populations and also the growth of these populations were examined monthly, some 10,000 specimens being involved. The young shrimps cannot survive in fresh water. They can withstand salinities as low as 0.5 ‰ but only develop normally in salinities ranging from 5 to 34 ‰. Their food is chiefly of animal origin (small fish, crustaceans, and polychaetes).
After a few months in brackish water, the shrimps migrate seawards and are fished chiefly over muddy bottoms at depths of 40 to 55 m.
At the various stages (post-larval, juvenile, adult) the Penaeus duorarum of Dahomey are slightly larger in size than the Florida Penaeus at similar stages.
INVESTIGACIONES SOBRE EL CICLO BIOLOGICO DEL CAMARON Penaeus duorarum, BURKENROAD 1939 EN DAHOMEY
Extracto
Este estudio constituye la primera investigación sobre el ciclo del camarón Penaeus duorarum Burkenroad, en Africa Ecuatorial (Dahomey); esta investigación se realizó desde octubre de 1962 hasta abril de 1964.
Las postlarvas emigran desde el mar hacia las aguas salobres de las lagunas. Se estudia mensualmente la relación que existe entre la salinidad y la densidad de cuatro poblaciones, así como su crecimiento, lo que supone el examen de 10.000 P. duorarum. Los camarones jóvenes no pueden vivir en agua dulce; soportan salinidades bajas de 0,5‰ pero sólo se crían normalmente con grados de salinidad de 5 a 34‰. La alimentación es generalmente de origen animal (pequeñnos peces, crustáceos, poliquetos).
Después de pasar algunos meses en aguas salobres, los camarones salen al mar; aquí se les pesca, principalmente en fondos cenagosos de 40 a 55 metros.
En las diversas fases (postlarvas, camarones jóvenes, camarones adultos), camarón Penaeus duorarum, de Dahomey, es algo más grande que los de Florida.
Le Décapode Macroure Natantia, Penaeus duorarum Burkenroad 1939, est répandu à la fois sur les côtes atlantiques de l'Amérique et de l'Afrique. C'est Burkenroad (1939) qui constata que l'espèce américaine, P. brasiliensis Latreille, correspondait en fait à trois espèces bien définies : P. brasiliensis Latreille, P. aztecus Ives et P. duorarum Burkenroad. Seule l'espèce P. duorarum existe sur la côte occidentale africaine. Il n'y a pas plus de différences entre P. duorarum d'Afrique et d'Amérique qu'entre les divers types de cette même espèce en Amérique (Holthuis, communication personnelle).
Sur la côte africaine, l'aire d'extension connue de P. duorarum est située entre la Mauritanie (Cap Blanc) et l'Angola (Luanda), c'est-à-dire entre le 21° de latitude nord et le 8° de latitude sud (Holthuis, 1952).
Dans ces mêmes eaux africaines, il existe également une autre espèce du même genre, Penaeus kerathurus Forskal 1775, mais celle-ci est absente de la côte atlantique américaine. Cette espèce est plus connue sous les noms de P. camarote Risso et de P. trisulcatus Leach; en Atlantique, elle s'étend du sud de l'Angleterre à l'Angola et est également commune en Méditerranée (Holthuis, 1952).
Il est facile de différencier ces deux espèces sur les côtes africaines par 4 caractères. Chez P. duorarum, le sixième somite abdominal montre, de chaque côté de la carène dorsale de ce somite, un sillon très net; il n'y a aucune trace de sillon chez P. kerathurus. D'autre part, P. duorarum est caractérisé par les sillons qui bordent latéralement la carène dorsale céphalothoracique sans atteindre l'extrémité postérieure de celle-ci; ces sillons atteignent nettement cette extrémité postérieure chez P. kerathurus et sont plus profondément creusés que chez P. duorarum. De plus, P. kerathurus porte des épines sur les coxae des trois premières paires de péréiopodes, ce qui n'est pas le cas chez P. duorarum. Enfin, l'abdomen des exemplaires de taille moyenne ou grande de P. kerathurus est zébré (bandes brunâtres alternant avec des bandes plus claires), alors que P. duorarum a un aspect uniforme.
Dans les eaux côtières dahoméennes, il existe également un autre Penaeidé très abondant, Parapenaeopsis atlantica. Cette espèce ne possède pas de sillon sur les côtés de la carène dorsale céphalothoracique; elle n'est connue que le long de la côte d'Afrique occidentale, entre le Ghana et l'Angola.
La présente étude est limitée à l'espèce Penaeus duorarum; elle correspond à des recherches effectuées dans la région de Cotonou (Dahomey) et en particulier dans une lagune saumâtre (lac Nokoué) (Fig. 1) d'octobre 1962 à mai 1964 (Hoestlandt, 1963, 1964, 1967) 1. Dans ce lac, P. kerathurus jeune n'est que très rarement capturé; nous n'y avons jamais pêché de P. atlantica.
En Amérique, P. duorarum se reproduit en mer, les jeunes larves remontent dans les estuaires ou en lagune; c'est en milieu saumâtre que se poursuit la croissance juvénile, et c'est en mer que les crevettes deviennent adultes.
Il n'existe, à notre connaissance, qu'une seule étude antérieure sur P. duorarum au Dahomey; il s'agit d'une note de 1908 (Lefebvre, 1908) qui souligne l'importance économique de cette crevette dans le lac Ahémé (Fig. 6) à une époque où P. duorarum était peu connue dans le lac Nokoué. Il faut également indiquer une étude récente (Monod, 1967) sur l'importance des diverses crevettes le long de la côte occidentale d'Afrique.
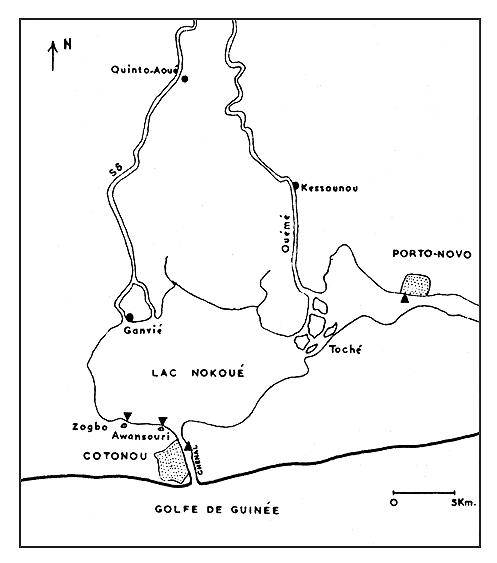
Fig. 1 Carte hydrographique de la Région du Lac Nokoué. Les triangles noirs indiquent les quatre stations de pêche de P. duorarum.
Au début de cette étude, il est intéressant d'indiquer sommairement les caractéristiques de l'Hydrographie et de la Climatologie du Dahomey; ceci permet de comprendre les variations de salinité de la région lagunaire car la Biologie de P. duorarum est en relation avec ces variations.
2.1 Eaux intérieures
2.1.1 Hydrographie. Une grande partie des eaux de ruissellement du Dahomey descend vers la mer sur de vastes pénéplaines granito-gneissiques ou sédimentaires. Ces eaux se concentrent dans les trois principaux fleuves, le Mono, le Couffo et l' Ouémé. Le dernier de ces fleuves nous intéresse plus spécialement car il aboutit dans la lagune de Porto-Novo et dans le lac Nokoué par le Toché; de plus, ces deux lagunes sont reliées entre elles par de nombreux bras et chenaux. Il faut ajouter qu'un fleuve beaucoup moins important, la Sô, se jette également dans le lac Nokoué (Fig. 1).
Le Bas Ouémé coule sur des alluvions récentes qui constituent une zone deltaïque de 75 km de longueur et de 10 à 15 km de largeur. De nombreux bras relient l'Ouémé à son affluent, le Zou, ainsi qu'avec la Sô. Dans cette zone deltaïque, la pente est très faible et la partie inférieure de cette zone est largement inondée en période de crue.
2.1.2 Importance des pluies. Les années correspondant à cette étude (1962–1964) sont des années pluvieuses; notre description climatologique (ainsi que l'étude biologique correspondante) ne concerneront donc pas le cas des années dites “sèches”.
Dans le Haut Dahomey, le régime climatique est continental de type soudanien avec une saison sèche de novembre à mars et une saison pluvieuse d'avril à novembre.
Dans le Moyen Dahomey, le régime est transitoire avec une diminution des précipitations en août.
Dans le Bas Dahomey, le régime comporte nettement deux saisons sèches (novembre à fin mars et août) ainsi que deux saisons pluvieuses (avril à juillet et septembre à octobre).
Le régime hydrologique de la lagune de Porto-Novo et du lac Nokoué est sous la dépendance de la première saison des pluies du Bas-Dahomey (avril à juillet) et surtout de la crue de l'Ouémé provoquée par les pluies du Haut Dahomey et du Moyen Dahomey. La crue de ces deux lagunes a lieu de juillet à fin novembre, avec un maximum généralement en septembre. C'est en mars et en avril que le débit de l'Ouémé est minimum.
Le lac Nokoué est peu profond (1 m par basses eaux), mais en période de crue, la hauteur d'eau du lac peut atteindre 2 m; dans ce cas, le courant de marée n'existe plus dans le chenal de Cotonou (Fig. 1).
2.1.3 Salinité.
- Lac Nokoué. En période de forte crue, de juillet ou août au début de novembre, les eaux du lac Nokoué sont douces et descendent vers la mer. Lors de la décrue de novembre, la marée amène de l'eau salée dans le chenal et la salinité augmente progressivement dans la lagune pour atteindre un maximum en février ou mars (salinité de 32 à 34%). L'action des pluies est ressentie dès les mois d'avril ou mai et la salinité de la lagune diminue assez rapidement.
- Lagune de Porto-Novo. Comme cette lagune est plus éloignée de la mer que le lac Nokoué, la salinité apparaîtra plus tardivement et n'atteindra pas de taux élevé. Cette lagune demeure douce jusqu'en janvier et la salinité maximum correspond au mois d'avril; elle ne dépasse pas 17‰.
- Ouémé et Sô. En mars et en avril, l'eau saumâtre remonte le cours inférieur de ces fleuves dans le Bas Dahomey jusqu'à 30 km en amont de la lagune de Porto-Novo pour l'Ouémé (Kessounou) et du lac Nokoué pour la Sô (Quinto-Aoué) (Fig. 1).
2.1.4 Température. Les lagunes côtières du Bas Dahomey ont une température équatoriale qui ne subit que très peu de variations au cours de l'année.
Dans le lac Nokoué, les températures de l'eau ont été régulièrement enregistrées. Le maximum de température se situe en avril (30, 3°C) et le minimum en août (27,2°C); la différence des températures au cours de l'année ne dépasse donc pas 3°C. Ces variations de températures annuelles sont trop faibles pour influencer la biologie des crevettes, alors qu'aux Etats-Unis elles jouent peut-être un certain rôle (Chin, 1961).
2.2 Eaux marines du plateau continental
Des recherches récentes (Crosnier et Berrit, 1966) nous apportent les éléments nécessaires à notre étude. La pente du fond marin côtier est assez faible jusqu'à 80 m pour devenir forte ensuite. Les fonds sont sableux jusqu'à 15 m, ils sont sablo-vaseux de 15 à 40 m, vaseux de 40 à 55 m et rocheux coralliens de 55 à 80 m.
Il existe une stratification des eaux marines côtières du Dahomey avec variations saisonnières liées au régime austral d'une période chaude centrée en février et d'une période froide centrée en août. On peut cependant distinguer nettement trois couches d'eau, malgré les modifications saisonnières.
Il y a d'abord une couche d'eaux chaudes dont la profondeur atteint de 20 à 30 m. La salinité varie de 32‰ à 35‰, mais elle peut descendre parfois à 20‰ aux abords de la côte. Les changements de salinité sont liés aux apports des eaux douces en période de pluies ou aux remontées des eaux salées profondes (“upwellings”). La température de ces eaux s'échelonne entre 25 et 29°C.
Il y a ensuite une couche de discontinuité ou couche de la thermocline qui a une épaisseur de 15 à 20 m et est localisée par conséquent entre 30 et 50 m. La salinité de cette couche varie peu (35,6 à 35,9‰) et la température est de l'ordre de 25°C.
Il y a enfin une couche profonde froide, au-dessous de 50 m. Dans cette couche, la salinité décroit de 0,004‰ par mètre et la température diminue de 0,03°C par mètre. A la profondeur de 80 m la salinité est de 35,7‰ et la température de 17°C.
Les stades larvaires et post-larvaires des Penaeides sont aisément caractérisés grâce aux recherches de Gurney (1942); il est très facile d'identifier P. duorarum grâce aux travaux de Williams (1959) et surtout de Dobkin (1961).
3.1 Technique
Les larves et les post-larves sont planctoniques, c'est pourquoi nous les recherchons avec des filets planctoniques de mailles différentes afin de capturer, soit les stades nauplius et protozoé, soit les stades mysis et post-larvaires.
Des pêches sont réalisées en mer lors d'une sortie à bord d'un chalutier, d'autres sont effectuées régulièrement dans le chenal ainsi que dans le lac Nokoué et à Porto-Novo (Fig. 1)
La longueur des crevettes est toujours exprimée en longueur du céphalothorax, mesurée entre le creux orbitaire droit et l'extrémité distale médiane du céphalothorax.
3.2 Résultats
Les stades protozoé ou mysis ne sont trouvés ni en mer ni en lagune. Aux Etats-Unis, ces stades ne sont que rarement capturés, et ceci toujours en mer (Idyll, 1964).
Les jeunes stades post-larvaires sont capturés dans le lac Nokoué. Le premier stade post-larvaire, qui succède au troisième stade mysis, est reconnaissable par la présence d'une seule épine dorsale sur le rostre; ce stade n'a pas été capturé. Le stade post-larvaire le plus jeune, pris dans la lagune, possède 4 épines rostrales: ce stade a été pris en onze exemplaires à l'amont du chenal (Fig. 1), en face d'Awansouri et même en face de Zogbo (un seul exemplaire en cette dernière station). Aux Etats-Unis, en Floride, c'est le même stade à 4 épines qui est le plus jeune stade pris en estuaire (Tabb, Dubrow and Jones, 1962).
La longueur de ce stade, exprimée en longueur du céphalothorax et mesurée au microscope, est en moyenne de 1,88 mm, ce qui correspond à une longueur totale de 7,5 à 8 mm. Signalons que ce stade ne mesure que 1,7 mm (longueur du céphalothorax) en Floride. Les post-larves du Dahomey sont donc légèrement plus grandes que celles de la Floride. Nous retrouverons ce caractère de dimensions plus grandes au cours de la croissance des P. duorarum du Dahomey.
Lebour (1959) donne le dessin d'une post-larve de P. duorarum, capturée en mer à hauteur du sud du Cameroun (2° 55'N. 9° 21'E.) le 28 novembre 1946, (2,5 mm de longueur totale de la carapace). Le dessin montre que cet exemplaire ne possède que 4 épines rostrales mais qu'il est déjà porteur de pléopodes biramés. Les exemplaires similaires que nous avons capturés au Dahomey ne comportent que des pléopodes uniramés Idyll (1964) reprenant l'étude de Lebour (1959), suggère qu'il y a peut-être une espèce ou une sous-espèce ouest-africaine différente de celle du sud de la Floride. Il n'y a pas lieu de retenir cette hypothèse mais plutôt d'estimer que la post-larve dessinée par Lebour (1959) était un spécimen exceptionnel anormal.
Le stade post-larvaire suivant comporte 5 épines rostrales; il n'est pas capturé dans le chenal, mais seulement à Awansouri et à Zogbo (3 exemplaires); il mesure 2,1 mm de longueur céphalothoracique. Les stades à 6 et 7 épines rostrales ne sont recueillis qu'à Zogbo (3 exemplaires).
Aucun stade post-larvaire n'a été pris à Porto-Novo.
Ces divers stades post-larvaires sont recueillis au Dahomey dès le mois de février. En Caroline, ils ne sont recueillis qu'à partir de mai; en Floride, on les prend dès le mois de mars; il n'est donc pas étonnant qu'on les capture plus tôt au Dahomey où les eaux sont plus chaudes.
En conclusion, les jeunes stades post-larvaires (4 épines rostrales) quittent la mer pour pénétrer dans le chenal de Cotonou. A ce stade, ces post-larves ont une nage relativement rapide; aidées par le courant, elles remontent facilement jusqu'à Zogbo situé à environ 8 kilomètres de la mer.
De 1962 à 1964, le lac Nokoué a été constamment en communication avec la mer par le chenal de Cotonou. Les eaux de cette lagune sont salées de décembre à juin et douces d'août à novembre. Lors de la présence d'eaux douces, il n'y a plus de crevettes, sauf à l'état sporadique, dans des zones dépourvues de courant (Zogbo, Awansouri) où la salinité descend à 0,5 ‰. Dans le cours inférieur de l'Ouémé et de la Sô, nous avons constaté que les crevettes suivent les eaux saumâtres jusqu'à leur limite amont (30 km de la mer) durant la période sèche (mars et avril). C'est ainsi qu'en avril 1964, nous avons constaté que la limite des eaux saumâtres atteignait Kessounou sur l'Ouémé (0,64‰) et Quinto-Aoué sur la Sô (3,21‰) (Fig. 1).
4.1 Croissance
4.1.1 Technique. Des échantillonnages de populations sont effectués mensuellement en quatre stations (Fig. 1), afin d'étudier la relation entre la salinité et la densité des populations et aussi afin de suivre la croissance des crevettes. La première station, celle dont les eaux sont relativement les moins salées, est située dans la lagune de Porto-Novo. Cette lagune est reliée au lac Nokoué par divers bras, en particulier par le Toché. Les crevettes qui s'y trouvent ne proviennent vraisemblablement que du lac Nokoué, car la distance entre Porto-Novo et Lagos (100 km) est trop importante pour que les crevettes arrivent habituellement par cette seconde voie lagunaire. Deux stations sont situées dans la lagune de Nokoué en une région réputée pour sa richesse en P. duorarum: Zogbo et Awansouri. Awansouri subit assez directement l'influence des eaux du chenal de Cotonou; Zogbo est une station qui ne subit guère cette influence. La quatrième station est située dans le chenal de Cotonou afin d'étudier plus particulièrement les mouvements de descente des crevettes vers la mer.
Nous avions l'intention d'utiliser un chalut à molettes tel que ceux qui sont employés sur les côtes d'Europe pour la pêche des crevettes côtières (Crangon). La différence très nette entre la biologie des Crangon et celle de P. duorarum a voué cet essai à un écheo. En effet, d'une part, les P. duorarum s'ensablent plus nettement que les Crangon durant la journée; d'autre part, les P. duorarum nagent la nuit. C'est pourquoi toutes les pêches de cette étude sont réalisées la nuit.
L'excellente connaissance pratique de l'écologie des P. duorarum lagunaires par les pêcheurs dahoméens nous a conduits à utiliser deux de leurs engine de pêche : l'“Azui” et le “Dohouihoué”.
l'Azui est un filet de forme pyramidale qui est traîné par deux pêcheurs. Nous utilisons un Azui de 3,50 m de longueur, dont l'ouverture trapézoïdale est de 1,60 m de hauteur, de 3 m de largeur à la base et de 2,50 m de largeur au sommet; les mailles de ce filet diminuent progressivement de l'ouverture (mailles de 2,5 cm) au fond de la poche (mailles de 0,5 cm).
Le Dohouihoué est un filet similaire disposé dans le courant et maintenu ouvert entre deux piquets plantés dans le sable du fond. Nous utilisons un Dohouihoué de 7 m de longueur; il possède une ouverture rectangulaire de 1,80 m de hauteur et de 4 m de largeur. Les mailles de ce filet mesurent 2,7 cm à l'ouverture et 0,6 cm au fond de la poche.
Afin de pouvoir comparer les diverses pêches entre elles, l'Azui est traîné entre 4 piquets de façon à parcourir 500 m à chaque circuit; généralement, il est effectué 4 tours, soit 2 000 m de parcours, afin d'obtenir un échantillonnage satisfaisant de la population. Le Dohouihoué est toujours monté dans le chenal pendant le reflux au début de la nuit, durant 4 heures consécutives.
Dans les quatre stations étudiées, il a été mesuré un total de 7.673 P. duorarum pour les prélèvements mensuels de décembre 1962 à avril 1964. L'échantillonnage le plus élevé est de 619 individus. Il a été capturé 931 crevettes à Porto-Novo en 7 échantillonnages, 2.225 à Zogbo en 11 échantillonnages, 2.776 à Awansouri en 12 échantillonnages et 1.741 dans le chenal de Cotonou en 10 échantillonnages. Il faut noter que les chiffres des prélèvements inférieurs à 30 P. duorarum ne sont retenus que pour l'examen de la densité des quatre populations étudiées.
Chaque crevette est mesurée à l'aide d'un pied à coulisse dont la précision est de 1/50 de mm; en fait, l'erreur absolue des mesures effectuées est égale à 1/10 de mm.
La longueur de chaque crevette est mesurée sur le céphalothorax entre la base du creux orbitaire droit et le milieu de la partie distale posterieure de la carapace.
La relation entre cette longueur du céphalothorax et la longueur totale de la crevette a été étudiée par Iversen et Idyll (1960) et l'on peut dire que la longueur du céphalothorax est égale au quart de la longueur totale, sans tenir compte de la légère différence de cette relation entre les mâles et les femelles.
Enfin, dans cette étude de la croissance, il n'est pas tenu compte de la différence de croissance entre les mâles et les femelles, car il est difficile de distinguer le sexe des jeunes crevettes, le petasma des mâles et le thelycum des femelles ne sont pas encore suffisamment différenciés.
4.1.2 Résultats
4.1.2.1 Densité de population et salinité. Pour chaque prélèvement dans les quatre stations étudiées, le Tableau I donne le total des crevettes capturées par unité de surface (parcours de l'azui ramené à 1 000 m pour Porto-Novo, Zogbo et Awansouri) ou par pêche au dohouihoué durant 4 heures dans le chenal de Cotonou. Le second chiffre entre parentèse exprime la salinité ‰ au moment du prélèvement.
Les Fig. 2 et 3 représentent graphiquement les salinités et les densités de populations pour chacune des quatre stations étudiées. De l'examen de ces graphiques, il ressort que la densité de population est en relation avec la salinité, en ce sens que les crevettes disparaissent lorsque les eaux deviennent douces; ainsi à Porto-Novo, il n'y a pas de P. duorarum si la salinité est inférieure à 0,5‰. Dans des eaux de faible salinité (0,5 à 1‰) il peut y avoir des crevettes (Awansouri et Zogbo, août à novembre 1963), mais la densité est faible. C'est à partir d'une salinité de 5‰ que les crevettes peuvent être abondantes (Porto-Novo, 19 février 1963). Enfin, c'est généralement au moment des fortes salinités que la densité des populations est maximale, en particulier à Zogbo et à Awansouri.
Si l'on compare les graphiques des 4 stations, on constate qu'à Porto-Novo P. duorarum est nettement saisonnière, alors qu'elle est capturée durant la plus grande partie de l'année à Zogbo et à Awansouri. Quant au chenal de Cotonou, le mouvement de descente des crevettes vers la mer est moins directement lié aux salinités que dans les stations lagunaires.
Il faut signaler enfin que les populations de 1964 sont, dans leur ensemble, moins denses que celles de 1963.
On peut conclure que les Penaeus en cours de croissance vivent aisément dans une eau de salinité minimum de 1‰ mais qu'elles ne sont abondantes qu'en eau de salinité égale ou supérieure à 5‰ et cela jusqu'à une salinité comparable à celle de l'eau de mer.
4.1.2.2. Croissance. On peut estimer que la méthode rigoureuse des prélèvements de crevettes (azui ou dohouihoué) permet un échantillonnage satisfaisant de la population au lieu de la pêche et que les différents prélèvements sont donc comparables entre eux.
L'étude graphique de la croissance donne lieu à la constitution d'histogrammes pour chacun des prélèvements mensuels dans les quatre stations. C'est dans ce but qu'à la suite de leur mensuration, les crevettes sont groupées par classes de 2 en 2 mm; chaque classe est figurée par son pourcentage afin de permettre la comparaison entre les histogrammes. Pour chaque prélèvement, nous avons établi le “mode” et la “moyenne arithmétique”. Les Fig. 4 et 5 donnent les graphiques du mode et de la moyenne de l'ensemble des prélèvements par station. Il faut rappeler que la moyenne arithmetique est généralement plus représentative de la croissance que le mode. Nous rappelons qu'il n'a pas été tenu compte des échantillonnages inférieurs à 30 individus.
TABLEAU I
Densités de populations et salinités
| Date | Porto-Novo | Zogbo | Awansouri | Chenal | ||||
| 5.12.62 | 28 | (8,42) | ||||||
| 7. 1.63 | 82 | (21,7) | ||||||
| 11. 1 | 48 | (22,7) | ||||||
| 25. 1 | 0 | (0,49) | ||||||
| 14. 2 | 154 | (29,20) | ||||||
| 19. 2 | 314 | (5,9) | ||||||
| 22. 2 | 98 | (29,2) | ||||||
| 1. 3 | 269 | (32) | ||||||
| 20. 3 | 250 | (9,3) | ||||||
| 22. 3 | 149 | (33,2) | ||||||
| 27. 3 | 413 | (33,95) | ||||||
| 29. 3 | 363 | (32,95) | ||||||
| 11. 4 | 44 | (30,55) | ||||||
| 12. 4 | 260 | (29,29) | ||||||
| 16. 4 | 130 | (27,47) | ||||||
| 23. 4 | 136 | (10,95) | ||||||
| 16. 5 | 133 | (23,48) | ||||||
| 21. 5 | 50 | (30,08) | ||||||
| 24. 5 | 78 | (25,77) | ||||||
| 30. 5 | 112 | (6,0) | ||||||
| 14. 6 | 51 | (22,70) | ||||||
| 20. 6 | 34 | (23,06) | ||||||
| 25. 6 | 60 | (19,33) | ||||||
| 27. 6 | 0 | (0,18) | ||||||
| 17. 7 | 46 | (9,92) | ||||||
| 19. 7 | 98 | (6,02) | ||||||
| 25. 7 | 101 | (5,57) | ||||||
| 30. 7 | 0 | (0,18) | ||||||
| 14. 8 | 38 | (1,0) | ||||||
| 16. 8 | 17 | (1,0) | ||||||
| 26. 8 | 51 | (0,98) | ||||||
| 16. 9 | 0 | (0,18) | ||||||
| 17. 9 | 13 | (0,57) | ||||||
| 18. 9 | 75 | (0,57) | ||||||
| 21.10 | 2 | (0,12) | ||||||
| 23.10 | 6 | (0,57) | ||||||
| 4.11 | 3 | (0,57) | ||||||
| 13.11 | 0 | (0,25) | ||||||
| 15.11 | 0 | (0,12) | ||||||
| 18.11 | 15 | (0,83) | ||||||
| 28.11 | 7 | (0,96) | ||||||
| 13.12 | 0 | (32,61) | ||||||
| 18.12 | 29 | (16,31) | ||||||
| 23.12 | 0 | (0,18) | ||||||
| 27.12 | 96 | (22,88) | ||||||
| 8. 1.64 | 0 | (0,75) | ||||||
| 13. 1. | 70 | (23,35) | ||||||
| 15. 1 | 97 | (20,72) | ||||||
| 21. 1 | 124 | (25,77) | ||||||
| 10. 2 | 62 | (31,98) | ||||||
| 13. 2 | 20 | (2,36) | ||||||
| 19. 2 | 81 | (32,74) | ||||||
| 24. 2 | 65 | (34,20) | ||||||
| 12. 3 | 407 | (34,85) | ||||||
| 13. 3 | 34 | (11,71) | ||||||
| 19. 3 | 15 | (34,20) | ||||||
| 24. 3 | 97 | (33,57) | ||||||
| 10. 4 | 554 | (34,02) | ||||||
| 16. 4 | 22 | (10,37) | ||||||
| 20. 4 | 67 | (29,42) | ||||||
| 24. 4 | 194 | (30,18) | ||||||
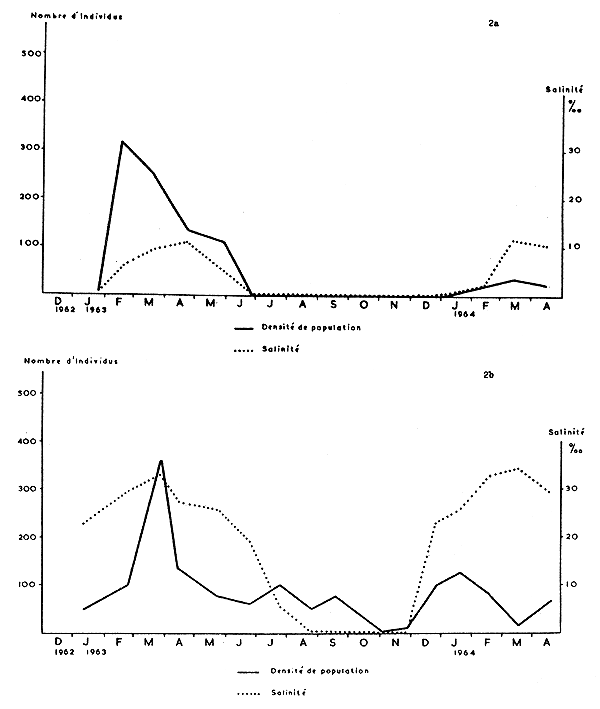
Fig. 2a Graphique des variations de la salinité de l'eau et de la densité de population des Penaeus duorarum à Porto-Novo.
2b A Zogbo.
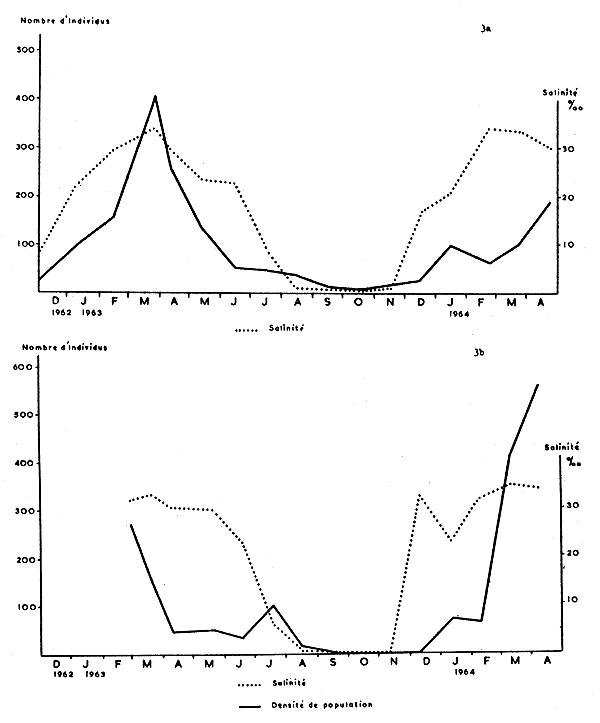
Fig. 3a Graphique des variations de la salinité de l'eau et de la densité de population des Penaeus duorarum à Awansouri.
3b Dans le chenal de Cotonou.
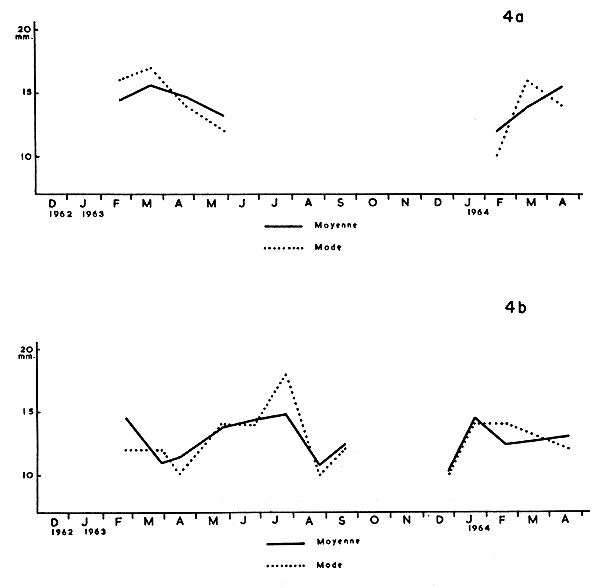
Fig. 4a Graphique des variations du mode et de la moyenne à Porto-Novo.
4b A Zogbo.
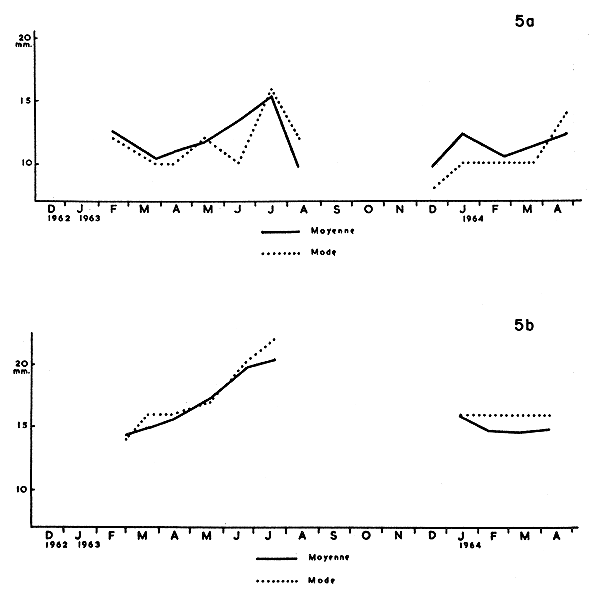
Fig. 5a Graphique des variations du mode et de la moyenne à Awansouri.
5b Dans le chenal de Cotonou.
L'examen de ces deux graphiques ne permet pas de caractériser la croissance des diverses populations. Ceci se comprend aisément pour le chenal de Cotonou où l'on ne capture que des crevettes relativement grandes en cours de descente vers la mer; il faut remarquer que la moyenne varie de 14,47 mm (février 1963) à 20,73 mm (juillet 1963) ce qui prouve que la descente ne correspond pas à une dimension précise, mais que les crevettes mesurent généralement de 15 à 20 mm de longueur du céphalothorax.
L'examen des stations lagunaires ne permet pas de suivre nettement la croissance des crevettes durant leur séjour en eau saumâtre. En effet, durant toute la phase saumâtre, il entre constamment de jeunes crevettes dans la lagune. Cependant, la population d'Awansouri présente la croissance moyenne (moyenne arithmétique) la plus importante entre les 27 mars et 17 juillet 1963 (16 semaines), soit 5 mm de longueur du céphalothorax. Cet accroissement correspond à 20 mm d'allongement total du corps; l'allongement moyen total serait donc de 1,3 mm par semaine. Or, selon Kutkuhn (1966), l'allongement de P. duorarum en Floride est de 3,4 mm, ce qui a été constaté avec rigueur sur des crevettes marquées. Mais cette différence entre les crevettes du Dahomey et celles de Floride ne peut être significative, car il ne s'agit au Dahomey, que de croissance moyenne d'une population dont les individus se renouvellent partiellement au cours des mois successifs.
Il est intéressant de rappeler que les crevettes les plus grandes, capturées en lagune ou dans le chenal, atteignaient une longueur céphalothoracique de 28 mm alors qu'aux Etats-Unis d'Amérique, en Floride (Tabb, Dubrow and Jones, 1962), elles ne dépassent pas la longueur de 20 à 22 mm. Nous retrouvons donc ici ce qui avait déjà été constaté pour les post-larves. Les P. duorarum du Dahomey sont donc un peu plus grandes que celles de la Floride. Est-ce dû à un facteur somatique ou génétique ?
4.2 Etat des glandes génitales
Les glandes génitales des femelles les plus grandes du lac Nokoué ont été disséquées et examinées au microscope. Il s'agit de femelles mesurant de 22 à 28 mm de longueur céphalothoracique, ce qui correspond à une longueur totale d'environ 10 cm.
Les ovaires de ces femelles sont transparents; les ovules sont clairs, apparemment dépourvus de vitellus et leur diamètre ne dépasse pas 41,5 μ. Il est bien évident que ces femelles sont nettement juvéniles et encore très éloignées de leur maturité; elles sont au stade “D” de King (1948), c'est-à-dire que les ovaires peu développés ont une teinte laiteuse; aucun stade “E” (ovaires jaunâtres) n'a été pris en lagune.
4.3 Alimentation
Un important travail de Hall (1962) relate l'examen des contenus stomacaux de plusieurs espèces de Penaeides de la Malaisie. L'alimentation de ces Penaeides est principalement animale (Polychètes, Mollusques, Crustacés, Poissons) et plus ou moins végétale. Eldred et al. (1961) constatent, à la suite d'autres auteurs, que P. duorarum est omnivore : les débris stomacaux sont principalement animaux (Polychètes, Crustacés, Mollusques, etc.) mais comportent également des végétaux (feuilles, rhizomes, algues).
Au Dahomey, nous avons examiné les contenus stomacaux de 20 P. duorarum mesurant de 12 à 18 mm de longueur du céphalothorax, prélevés dans l'échantillonnage de la population d'Awansouri (12 avril 1963).
Le Tableau II présente les principaux éléments des contenus stomacaux. L'importance de l'aliment envisagé est figurée par des croix: + = peu abondant; ++ = abondant; +++ = très abondant.
TABLEAU II
Contenus stomacaux
| Numérotation des crevettes | Poissons | Crustacés benthiques | Crustacés planctoniques | Polychètes | Mollusques | Végétaux |
| 1 | ++ | ++ | + | |||
| 2 | ++ | ++ | + | |||
| 3 | ++ | +++ | ||||
| 4 | ++ | ++ | + | |||
| 5 | + | + | + | + | + | |
| 6 | +++ | + | + | |||
| 7 | ++ | + | + | + | ||
| 8 | + | ++ | + | + | + | |
| 9 | + | ++ | + | |||
| 10 | + | + | + | + | ||
| 11 | ++ | + | + | |||
| 12 | ++ | ++ | + | |||
| 13 | + | + | + | |||
| 14 | ++ | + | + | + | ||
| 15 | +++ | + | ||||
| 16 | ++ | + | + | |||
| 17 | ++ | + | ++ | |||
| 18 | ++ | + | ||||
| 19 | + | ++ | ||||
| 20 | ++ | ++ | + |
De l'examen de ce tableau, il ressort que les crevettes de cette population d'Awansouri se nourrissent principalement de poissons et de crustacés. Il s'agit de petits poissons dont les vertèbres mesurent de 0,25 à 0,35 mm de longueur; dans un seul estomac il est trouvé des écailles relativement grandes mesurant 2,55 mm de diamètre. Les crustacés sont très divers, principalement benthiques (Amphipodes, petits Décapodes) quelquefois planctoniques (Copépodes). Les Polychètes sont beaucoup moins fréquentes, ce qui est probablement lié à leur densité plus faible que celle des crustacés et des poissons dans la lagune. Il n'a été trouvé de mollusques que dans un seul estomac: il s'agit de Gastropodes. Les végétaux sont fréquemment représentés, mais toujours en faible quantité. Il y a d'autres débris difficilement identifiables (Chétognates, etc.).
En résumé, les P. duorarum de la lagune semblent être principalement carnivores avec un léger apport végétal.
4.4 Expérimentation
Quelques essais d'élevage sont réalisés, soit en aquarium de verre, soit en bacs plastiques. A cause de la fragilité des crevettes, celles-ci, pêchées la nuit à Awansouri dans les nasses, sont transportées en cuvettes jusqu'au laboratoire, car elles supportent mal le transport à sec.
Elevage en grand aquarium. Les crevettes sont normalement ensablées durant le jour; seuls les yeux sont visibles à la surface du sable; la nuit, les crevettes nagent dans l'aquarium. La nourriture est constituée par des poissons morts, divers crustacés et quelques débris végétaux; les crevettes semblent préférer les poissons aux crustacés. Ces crevettes vivent dans des conditions satisfaisantes puisque les mues, qui sont toujours un moment critique dans la vie des crustacés, s'effectuent normalement.
Elevage en petits aquariums. Dans le but de réaliser des élevages comparables, on utilise 7 petits aquariums de verre d'une contenance de 5 litres d'eau. Les divers aquariums contiennent des eaux de salinité croissante: 0,07‰; 2‰; 5‰; 10‰; 15‰; 20‰; 33‰. Les crevettes utilisées mesurent de 9 à 18 mm de longueur du céphalothorax et sont prélevées après un séjour de 24 heures dans le grand aquarium de 60 l d'eau (salinité 20‰). On dispose 5 crevettes par petit aquarium et l'expérience dure 5 jours, car la petite dimension des aquariums ne permet que des essais très limités dans le temps sur l'influence de la salinité.
En eau douce (salinité 0,07‰), les crevettes meurent après 2 à 6 heures d'expérimentation.
Dans une eau de salinité de 2‰, les crevettes meurent après 3 à 24 heures d'expérimentation.
Dans les eaux de salinité de 5‰, 10‰, 15‰ et 33‰, les crevettes vivent facilement durant les 5 jours de l'expérience. Il y a même des mues dans plusieurs aquariums, aussi bien dans les eaux de salinité de 5‰ que dans celles de 33‰.
Ces expériences permettent d'affirmer que les P. duorarum vivent aisément dans des eaux de salinité variant entre 5 et 33‰; elles ne peuvent subsister longtemps dans des eaux de salinité égale ou inférieure à 2‰, au moins dans les conditions de l'expérience, c'est-à-dire lorsque les crevettes sont prélevées dans une eau de salinité de 20‰ pour être portées directement dans les aquariums à salinité égale ou inférieure à 2‰.
En résumé, ces divers essais expérimentaux prouvent que P. duorarum ne peut vivre en eau douce. D'autre part, les crevettes mesurant plus de 9 mm de longueur du céphalothorax ne peuvent supporter le passage brusque d'une salinité de 20‰ à une salinité égale ou inférieure à 2‰.
Les pêches du 14 août 1963 à Awansouri (salinité 1‰) et du 26 août 1963 à Zogbo (salinité 0,98‰), prouvent que des P. duorarum mesurant en moyenne 11 mm de longueur du céphalothorax peuvent supporter pendant quelque temps cette faible salinité; mais l'abaissement de salinité s'est produit lentement dans ces deux stations au cours des mois de juillet et d'août. Il est également possible que certaines crevettes soient plus résistantes et que les autres crevettes soient descendues vers la mer ou aient péri en lagune.
Avant 1962, aucune capture de P. duorarum n'avait été signalée en mer au large de Cotonou. C'est pourquoi le problème de la reproduction de cette espèce en lagune s'est posé. Mais, comme cette espèce ne se reproduit qu'en mer sur les côtes atlantiques des Etats-Unis, il semblait difficile d'admettre un cycle biologique différent sur les côtes équatoriales africaines. D'autre part, les pêcheurs de la lagune de Cotonou reconnaissent qu'en cas de fermeture naturelle de la lagune pendant plusieurs années (comme cela s'est produit dans le passé), on ne pêche plus de P. duorarum dans la lagune, mais seulement des Macrobrachium.
5.1 Localisation
En octobre 1962, nous avons examiné les pêches du chalutier “Avrekété” au large de Cotonou. Le chalut est traîné sur des fonds de 12 à 17 mètres. Les Penaeides capturées sont des Parapenaeopsis atlantica abondantes sur ces fonds et de rares spécimens de Penaeus kerathurus; aucum P. duorarum n'a été capturé.
En 1963, le chalutier “Zéphir” a pêché par des fonds de 10 à 40 m; grâce à l'obligeance de M. Paraiso, Directeur du Service des pêches du Dahomey, et de la Direction de la SOCAP, nous avons examiné les pêches de ce chalutier de juin à fin août.
Par des fonds de 10 à 20 m (de 1 à 8 km au large), le chalutier ne prenait que des espèces de Penaeides pêchées antérieurement par l“Avrekété”. Par des fonds de 20 à 40 m (entre 8 et 18 km au large), le “Zéphir” a capturé des P. duorarum en nombre appréciable. Il s'agit de femelles plus grandes que celles de la lagune. Beaucoup d'entre elles mesurent de 40 à 50 mm de longueur du céphalothorax; la plus grande longueur observée à Cotonou est de 50,9 mm ce qui correspond à une crevette de longueur totale égale à 24 cm. Cette longueur est comparable à la plus grande longueur (50 mm) signalée par Iversen et Idyll (1960) sur les côtes de Floride. D'autre part, à Lagos (26 août 1963), nous avons examiné une femelle mesurant 52 mm de longueur du céphalothorax. Chez les adultes, comme c'est le cas pour les autres stades de croissance, les P. duorarum du Dahomey sont un peu plus grandes que celles des côtes de Floride.
Les résultats les plus intéressants de la localisation des P. duorarum en mer sont obtenus par les pêches expérimentales du navire océanographique de l'Office de la Recherche Scientifique et Technique d'Outre-Mer “Ombago” en octobre 1963 (Crosnier et Berrit, 1966). Le littoral du Dahomey et celui du Togo ont été sillonnés de la côte jusqu'à 20 km au large.
Parmi les 29 chalutages réalisés et inventoriés (Paraiso, 1964), 8 seulement contenaient des P. duorarum (Fig. 6, lettres A à H). La plupart de ces chalutages se situent dans la zone vaseuse de 40 à 55 m de profondeur (bande pointillée de la Fig. 6). Cette bande vaseuse est localisée entre la zone de sable coquillier grossier et la zone rocheuse coralliaire. D'autre part, elle correspond à la couche d'eaux marines située au-dessus de la thermocline.
On peut donc estimer qu'au large du Dahomey, les P. duorarum vivent principalement sur des fonds qui sont à 12 km de la côte et qui ne mesurent que de 2 à 5 km de largeur (Fig. 6). Au Nigéria, de Vriès et Lefevere (1966) signalent des captures de P. duorarum entre 18 et 45 m de profondeur.
Les P. duorarum pris au Dahomey par des fonds inférieurs (20 à 40 m) durant l'été de 1963 étaient toutes des femelles. Selon l'avis du patron du chalutier, il n'en a pas été pris sur ces fonds, ni en hiver, ni au printemps; il s'agit peut-être de femelles qui viennent pondre dans des eaux moins profondes. Cependant en mai 1964, un autre chalutier a pêché des P. duorarum lors des chalutages sur des fonds de 30 à 40 m; il s'agit de crevettes de diverses tailles, mais toujours plus grandes que les crevettes de la lagune.
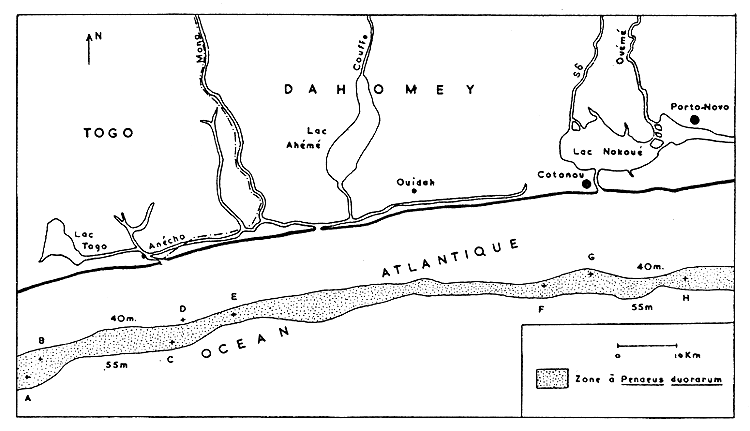
Fig. 6 Carte des fonds marins à Penaeus duorarum le long du littoral dahoméen.
D'autre part, nous avons examiné des documents non publiés dûs aux recherches de Longhurst et Mann. Le long des côtes de la Nigéria (entre Lagos et la Kulana River), il a été capturé, en 1963, des P. duorarum sur des fonds variant entre 15 et 60 m, alors que les chalutages ont été effectués entre 10 et 250 m de profondeur.
Sur les côtes américaines (Floride), les P. duorarum vivent principalement sur des fonds de 18 à 36 m; la plus grande profondeur observée est de 70 m (Idyll, 1964). Ces fonds sont constitués par de la vase avec 20% de sable coralliaire. Il est donc intéressant de souligner qu'au large du Dahomey, les P. duorarum vivent généralement plus profondément qu'en Floride.
5.2 Etat des glandes génitales
Les plus grandes femelles capturées au large de Cotonou en juin et juillet 1963 ainsi que celles examinées à Lagos (août 1963) possèdent des ovaires volumineux de couleur jaune verdâtre, c'est-à-dire entre les stades “Y” et “R” de King (1948); ce sont donc des ovaires proches de la maturité. Les plus grands ovules mesurent 3/10 de mm de diamètre; cette dimension correspond à celle des ovules mûrs des P. duorarum de Floride (Dobkin, 1961).
La pêche de P. duorarum est actuellement importante dans le lac Nokoué alors qu'elle est réduite dans le lac Ahémé (Fig. 6); c'est une situation inverse de celle du début de ce siècle (Lefebvre, 1908). Une estimation de la pêche annuelle par les nasses et les filets à crevette s'avère difficile. D'après un rapport du Service des Eaux et Forêts du Dahomey, 1957 à 1959, la productivité du lac Nokoué serait de plus de 1 000 tonnes de crevettes par an. Ces chiffres semblent élevés; on peut penser qu'un chiffre de 500 tonnes serait actuellement plus proche de la réalité. Ces crevettes sont vendues aux Dahoméens après avoir été séchées et fumées; une partie de ces crevettes est actuellement exportée vers la France. D'autre part, la pêche de ces crevettes en mer n'a commencé qu'en 1963.
En 1966, des relevés statistiques du Service Dahoméen des Pêches, signalent que les deux usines de congélation de Cotonou ont traité environ 60 tonnes de P. duorarum provenant principalement du lac Nokoué (46 t), puis du lac Ahémé (10 t), de Porto-Novo (2 t) et de l'Océan Atlantique (2 t). Ces crevettes sont expédiées en France après cuisson et congélation.
Burkenroad, M.D., 1939 Further observations on Penaeidae of the northern Gulf of Mexico. Bull.Bingham oceanogr.Coll., 6(6):1–62
Chin, E., 1961 A trawl study of an estuarine nursery area in Galveston Bay, with particular reference to Penaeid shrimp. University of Washington, Thesis, 113 p.
Crosnier, A. et G.R. Berrit, 1966 Fonds de pêche le long des côtes des Républiques du Dahomey et du Togo. Cah. O.R.S.T.O.M. Oceanogr., 4(1)suppl., 144 p.
de Vriès, J. et S. Lefevere, 1966 A maturity key for Penaeus duorarum Burkenroad 1939 of both sexes. Paper presented to FAO Fourth Regional Conference, Abidjan, November, 1966, 16 p.
Dobkin, S. 1961 Early development stages of pink shrimps, Penaeus duorarum, from Florida Waters. Fish.Bull.U.S. 61(190):321–49
Eldred, B. et al., 1961 Biological observations on the commercial shrimp, Penaeus duorarum Burkenroad, in Florida waters. Prof.Pap.Ser.mar.Lab.Fla, (3):139 p.
Gurney, R., 1942 Larvae of Decapod crustacea. London, Ray Society, 306 p.
Hall, D.N.F., 1962 Observations on the taxonomy and biology of some Indo-West-Pacific Penaeidae. (Crustacea Decapoda). Fish.Publs colon.Off., (17):229
Hoestlandt, H., 1963 Premières données sur la relation entre le cycle biologique et la salinité chez la crevette Penaeus duorarum Burkenroad du Lac Nokoué. Centre Technique Forestier Tropical, Rapport de Travaux d'Etudes sur la Pêche lagunaire au Dahomey. pp 72–96
Hoestlandt, H., 1964 Données complémentaires sur la relation entre le cycle biologique et la salinité chez la crevette Penaeus duorarum Burkenroad du Lac Nokoué. Centre Technique Forestier Tropical, Rapport de Travaux d'Etudes sur la Pêche lagunaire au Dahomey, pp 121–36
Hoestlandt, H., 1967 Premières recherches sur le cycle biologique de Penaeus duorarum Burkenroad 1939 en Afrique occidentale (Dahomey). Pubs scient.tech.Res.Commn Orgn.Afr., 96:475–98
Holthuis, L.B., 1952 Crustacés Décapodes, Macroures (Expédition Océanographique belge dans les eaux côtières africaines de l'Atlantique Sud). Bull.Inst.T.Sci.nat. Belg., 3(2):88 p.
Idyll, C.P., 1964 A summary of information on the Pink Shrimp, Penaeus duorarum. Paper presented to CSA Specialist Meeting on Crustaceans, Zanzibar, April 19–26, 1964, 22 p.
Iversen, E.S. and C.P. Idyll, 1960 Aspects of the biology of the Tortugas Pink Shrimp, Penaeus duorarum. Trans.Am.Fish.Soc., 89(1):1–8
King, J.E., 1948 A study of the reproductive organs of the common marine shrimp, Penaeus setiferus (Lin.). Biol.Bull.mar.biol.Lab., Woods Hole, 94(3):244–62
Kutkuhn, J.H., 1966 Dynamics of a penaeid shrimp population and management implications. Fishery Bull.Fish Wildl.Serv.U.S., 65(2):313–38
Lebour, M.V., 1959 The larval decapod crustacea of tropical West Africa. Atlantide Rep. (5):119–43
Lefebvre, M., 1908 Notice sur le Penaeus brasiliensis, crevette du Bas-Dahomey (crevette du lac Ahémé). Bull.Mus.Hist.nat., Paris, (14):267–70
Monod, Th., 1967 Crevettes et crabes de la côte occidentale d'Afrique. Pubs scient.tech.Res. Commn Orgn Afr., 96:103–234
Paraiso, F., 1964 Compte-rendu de campagne de chalutage du Navire océanographique “OMBANGO” Service des Pêches du Dahomey, 18 p.
Tabb, D.C., D.L. Durrow et A.E. Jones, 1962 Studies on the biology of the pink shrimp Penaeus duorarum Burkenroad, in Everglades National Park, Florida. Tech. Sor.Fla St.Bd Conserv., (37):5–30
Williams, A.B., 1959 Spotted and brown shrimp postlarvae (Penaeus) in North Carolina. Bull. Mar.Sci.Gulf Caribb., 9(3):281–90
![]()
![]()
![]()